La malaria y la deficiencia de hierro son importantes problemas de salud pública a nivel mundial, con una amplia superposición epidemiológica. Sin embargo, el impacto de la escasez de hierro en las células del hospedador, más allá de la anemia, sigue siendo incierto en el contexto de la malaria. Tras experimentar con ratones, los autores de este artículo científico arrojan luz sobre el papel del hierro en el complejo equilibrio entre protección y patología en la malaria.
Presentación
“Celular iron governs the host response to malaria” (“El hierro de las células regula la respuesta del hospedador a la malaria”), es un artículo científico elaborado por Wideman, S. K.; Frost, J. N.; Richter, F. C.; Naylor, C.; Lopes, J., M.; Viveiros, N.; Preston, A. E.; Teh, M. R.; White, N.; Yusuf, S.; Draper, S., J.; Armitage, A., E.; Duarte, T., L.; y Drakesnith, H., publicado en la revista científica PLOS Pathogens en Octubre de 2023, en el que sus autores arrojan luz sobre el papel del hierro en las células del hospedador más allá de la anemia en el contexto de la malaria, tras realizar una serie de experimentos con ratones.
La malaria es una enfermedad infecciosa grave y potencialmente letal que afecta a casi 250 millones de personas cada año en todo el mundo. Es causada por parásitos del género Plasmodium transmitidos entre humanos por mosquitos Anopheles.
El hierro, por su parte, es un micronutriente esencial que la mayoría de los organismos vivos necesitan para mantener procesos fisiológicos y bioquímicos, como el transporte y almacenamiento de oxígeno, el metabolismo celular y las reacciones de reducción-oxidación. A pesar de la importancia del hierro, su deficiencia es extremadamente común humanos, siendo especialmente frecuente en las zonas endémicas de malaria.
Existe una relación compleja e incompletamente comprendida entre el hierro y esta enfermedad. La anemia inducida por la deficiencia de hierro puede proteger al hospedador de la malaria al limitar el crecimiento del parásito. Por otro lado, la deficiencia de hierro puede alterar significativamente la función de las células inmunitarias como linfocitos y neutrófilos, que van a combatir la enfermedad. Sin embargo, se desconoce cómo afecta la deficiencia de hierro a otras células del hospedador en el contexto de la malaria. Además, aunque se sabe que la deficiencia de hierro del hospedador influye en la infección por malaria, los mecanismos que afectan la salud del mismo o la virulencia de Plasmodium siguen siendo en gran medida desconocidos.
Para abordar estas cuestiones, los autores de este trabajo utilizaron ratones genéticamente modificados con una menor captación celular de hierro, pero sin anemia, y los infectaron con una cepa murina de malaria.
Así, este estudio destaca el papel fundamental del estado del hierro en las células del hospedador en la malaria y puede tener implicaciones para el enfoque de tratamientos, tanto de la malaria como de la deficiencia de hierro, en las regiones endémicas de dicha enfermedad.
Materiales y Métodos
A continuación, se narran los materiales empleados y los métodos de trabajo seguidos por los autores de este estudio científico sobre el papel del hierro en las células del hospedador de malaria.
Ratones
Los ratones TfrcY20H/Y20H fueron proporcionados por el profesor Raif Geha, del Hospital Infantil de Boston/Facultad de Medicina de Harvard, y se criaron en la Universidad de Oxford.
Se adquirieron ratones de control de tipo silvestre C57BL/6JOlaHsd de Envigo y se alojaron junto con los ratones TfrcY20H/Y20H durante 2-3 semanas antes de la infección por P. chabaudi.
Todos los ratones se alojaron en jaulas individuales, ventiladas, libres de patógenos específicos y bajo condiciones normales de luz, y se alimentaron con pienso estándar que contenía 188 ppm de hierro ad libitum.
Para los experimentos usaron ratones hembra de 8 a 13 semanas de edad. Se utilizaron exclusivamente hembras para evitar la pérdida de animales por peleas y para minimizar el riesgo de eventos adversos graves por infección por P. chabaudi, el cual es mayor en machos. La eutanasia se realizó por asfixia mediante concentraciones crecientes de CO2, y la muerte se confirmó por dislocación cervical.

Parásitos e infección
Obtuvieron parásitos transgénicos de P. chabaudi chabaudi AS recientemente transmitidos por mosquitos y que expresan GFP, del Repositorio Europeo de Reactivos de Malaria de la Universidad de Edimburgo.
Para generar eritrocitos infectados (iRBC) para infecciones por P. chabaudi en la fase sanguínea, las reservas de parásitos congelados se descongelaron rápidamente a mano y se inyectaron por vía intraperitoneal (i.p.) en un ratón silvestre. Una vez que la parasitemia ascendente alcanzó el 0,5-2%, el animal fue eutanasiado y desangrado mediante punción cardíaca. Las infecciones experimentales posteriores se iniciaron inmediatamente a partir de la sangre recolectada, mediante la inyección intravenosa (i.v.) de 10⁵ eritrocitos infectados (iRBC) en 100µL de solución de Alsever. Los ratones control no infectados recibieron únicamente la solución de Alsever.
Para monitorizar la infección por P. chabaudi, recolectaron sangre mediante micromuestreo de la vena caudal de los ratones infectados. La parasitemia, el recuento de iRBC y el recuento de eritrocitos, se midieron mediante citometría de flujo. Para ello, se diluyeron 2µL de sangre en 500µL de solución de Alsever inmediatamente después de la recolección. La solución se diluyó posteriormente 1:10 en PBS antes de su análisis en un citómetro de flujo Attune NxT (Thermo Fisher Scientific). Se adquirió un volumen fijo de cada muestra, lo que permitió la cuantificación del total de eritrocitos e iRBC por µL de sangre.

Tratamiento con αBMP6
Para elevar experimentalmente los niveles de hierro sérico, se administró un anticuerpo monoclonal IgG humano bloqueador de αBMP6 que presenta reactividad cruzada con BMP6 murino.
Los ratones control recibieron un anticuerpo de control de isotipo IgG4 humano.
Ambos anticuerpos se diluyeron en 100μL de PBS y se inyectaron por vía intraperitoneal a una dosis aproximada de 10mg/kg de peso corporal.
Procesamiento de tejidos
Los órganos y tejidos se extrajeron poco después de la eutanasia y se mantuvieron refrigerados hasta su posterior análisis.
Los índices de hígado y bazo se calcularon como la masa de los órganos respectivos en relación con el peso corporal del ratón.
La sangre se recolectó en tubos de recolección de sangre adecuados, ya sea mediante muestreo de la vena caudal o mediante punción cardíaca después de la eutanasia.
El suero se preparó mediante centrifugación de los tubos de recolección a 10 000 x g durante 5 minutos y se almacenó a -80°C.
Análisis de sangre
El recuento de glóbulos rojos, la hemoglobina y el volumen corpuscular medio, se midieron a partir de sangre total utilizando un analizador hematológico automático KX-21N (Sysmex).
Los niveles séricos de ANG-2 y ALT se midieron según las instrucciones del fabricante, utilizando el kit ELISA para ALT de ratón y el kit ELISA Quantikine para angiopoyetina-2 de ratón/rata, respectivamente.
Las citocinas séricas se midieron mediante el inmunoensayo basado en microesferas LEGENDplex Mouse Inflammation Panel.

Ensayo de invasión in vitro por P. chabaudi
Para evaluar la susceptibilidad de los eritrocitos de tipo silvestre y TfrcY20H/Y20H a la invasión por P. chabaudi, recolectaron sangre de un ratón de tipo silvestre infectado con P. chabaudi durante la fase ascendente de la parasitemia (eritrocitos donantes/Y) y de ratones de tipo silvestre y TfrcY20H/Y20H no infectados (eritrocitos diana/X).
Para eliminar los leucocitos, filtraron la sangre a través de una columna de celulosa.
Los eritrocitos diana se marcaron fluorescentemente con 1μM de CellTrace Far Red en PBS, diluyendo la sangre 1:10 con la solución de CellTrace e incubando en la oscuridad durante 15 minutos a 37°C, mezclando las muestras cada 5 minutos.
Posteriormente, las células se lavaron dos veces en medio R10 (RPMI-1640 con 10% de FBS, 2mM de glutamina, 1% de penicilina-estreptomicina y 50μM de 2-mercaptoetanol) y se resuspendieron en medio R10 suplementado con 0,5mM de piruvato de sodio.
Sembraron 2 x 10⁷ eritrocitos donantes y 2 x 10⁷ eritrocitos diana marcados con fluorescencia en el mismo pocillo de una placa de 96 pocillos y se incubaron durante la noche (~16h) en un recipiente con vela a 37°C.
La invasión se midió como eritrocitos GFP+ y se comparó calculando el índice de susceptibilidad de cada tipo de eritrocito (X e Y).
Mediciones de hierro
Las mediciones de hierro sérico se realizaron en un analizador automatizado Abbott Architect c16000 por el personal de Bioquímica Clínica de los Hospitales Universitarios de Oxford, utilizando el kit MULTIGENT Iron (Abbott), o un analizador automatizado Pentra C400 con el kit Iron CP ABX Pentra (HORIBA Medical).
Para las mediciones de hierro hepático no hemínico se recolectaron fragmentos de tejido hepático, se congelaron rápidamente y se almacenaron a -80°C. El tejido se secó a 100°C durante aproximadamente 6 horas, se pesó y luego se digirió en ácido tricloroacético al 10% /ácido clorhídrico al 30% en agua durante aproximadamente 20 horas a 65°C.
Posteriormente, se añadió un reactivo cromogénico que contenía 0,1% de ácido batofenantrolinadisulfónico, 0,8% de ácido tioglicólico y 11% de acetato de sodio en agua, y se midió la absorbancia a 535nm.
El contenido de hierro se determinó comparando las muestras con una curva de calibración de citrato férrico amónico diluido en serie.

Citometría de flujo
Las suspensiones de células individuales para citometría de flujo se prepararon mediante disociación mecánica y enzimática.
Los bazos se filtraron a través de filtros celulares de 70μm, se incubaron con 120 U Kunitz/mL de desoxirribonucleasa I en medio R10 durante 15 min con agitación, y se filtraron a través de filtros celulares de 40μm.
Los hígados se perfundieron con PBS con 10% de FBS antes de su extracción. Para preparar las suspensiones de células individuales, los hígados se disgregaron con tijeras, se incubaron con 0,5mg/mL de colagenasa IV y 120 U Kunitz/mL de ADNasa I en R10 durante 45min con agitación, y se filtraron a través de filtros celulares de 70μm.
Posteriormente, se realizó la lisis de los eritrocitos, resuspendiendo las células centrifugadas en tampón de cloruro de amonio con Tris (0,017 M Tris / 0,14 M NH4Cl, ajustado a pH 7,2 con HCl) e incubándolas durante aproximadamente 5 minutos en hielo antes de lavarlas con R10.
Las células inmunitarias se aislaron del hígado mediante separación con Percoll. Las suspensiones de células individuales se colocaron suavemente sobre Percoll al 33% y se centrifugaron durante 25 minutos a 800 x g. Tras la centrifugación, se descartó el sobrenadante y los leucocitos restantes se lavaron dos veces con R10.
Para la tinción intracelular de citocinas, los esplenocitos se cultivaron ex vivo en medio R10 a una concentración de 5–2 x 10⁵ células/mL, en placas de 96 pocillos con fondo redondo tratadas para cultivo celular, con el inhibidor del transporte de proteínas Brefeldina A durante 4–6h a 37°C y 5% de CO₂. Para activar las células T, se añadió 0,5μg/mL de anticuerpo anti-CD3 de ratón a los esplenocitos de ratones infectados con P. chabaudi.
Las células se contaron utilizando un contador y analizador celular CASY (BOKE), y se tiñeron 1–5 x 10⁶ células para citometría de flujo. Para ello, las células se lavaron en PBS, se bloquearon con TruStain FcX y se tiñeron con un colorante de viabilidad o LIVE/DEAD durante ~10 min a 4°C en la oscuridad. A continuación, se añadieron anticuerpos conjugados con fluoróforos a las células y se incubaron durante ~20 min. Las células se lavaron dos veces en PBS y se fijaron incubándolas con Fixation Buffer durante ~10 min a 4°C en la oscuridad.
Como alternativa, las células se fijaron y permeabilizaron utilizando eBioscience FOXP3/Transcription Factor Staining Buffer Set, y se realizó la tinción del factor de transcripción, según las instrucciones del fabricante.
La tinción intracelular de citocinas se realizó tras la permeabilización con el tampón de lavado para tinción intracelular durante aproximadamente 30 minutos, siguiendo el protocolo del fabricante.
Finalmente, las muestras se analizaron en un citómetro de flujo Attune NxT o BD LSR Fortessa X-20 (BD).
Cultivo in vitro de células inmunitarias primarias
Las células T CD4+ vírgenes y las células B se aislaron siguiendo las instrucciones del fabricante, a partir de suspensiones mixtas de células individuales de esplenocitos y ganglios linfáticos, utilizando el kit de aislamiento de células T CD4+ vírgenes EasySep de ratón, o a partir de suspensiones de células individuales de esplenocitos, utilizando el kit de aislamiento de células B EasySep de ratón.
Las células aisladas se marcaron fluorescentemente con 5μM de CellTrace Violet en PBS durante 8 minutos a 37°C y se lavaron dos veces con medio R10. El recuento celular se realizó con un contador y analizador celular CASY.
Para las células T CD4+, recubrieron previamente placas de 96 pocillos de fondo plano para cultivo celular con 5μg/ml de anticuerpo anti-CD3 de ratón y sembraron las células a una concentración de 5 x 10⁵ células/ml.
Las células se cultivaron en un medio polarizador Th1 compuesto por R10 con 1μg/mL de anticuerpo anti-CD28 de ratón, 5μg/mL de anticuerpo anti-IL-4 de ratón, 10ng/mL de IL-12, 25 U/mL de IL-2 y 50μM de 2-mercaptoetanol. El medio se reemplazó después de 48 horas de cultivo. Para suplementar el medio de cultivo con hierro, se añadió sulfato ferroso heptahidratado.
Las células B se cultivaron a una concentración de 7,5 x 10⁵ células/mL en placas de 96 pocillos de fondo plano para cultivo celular, en medio R10 suplementado con 1% de aminoácidos MEM, 2μg/mL de LPS, 10ng/mL de IL-4, 10ng/mL de IL-5 y 50μM de 2-mercaptoetanol. Añadieron citrato férrico amónico en las concentraciones especificadas para suplementar el medio con hierro.
Las células T CD4+ se cultivaron durante 96h y las células B durante 72h, ambas a 37°C y 5% de CO₂, antes de la tinción para citometría de flujo.
Análisis de la expresión génica
El análisis de la expresión génica mediante PCR cuantitativa en tiempo real se realizó en muestras de hígado conservadas en solución estabilizadora RNAlater y almacenadas a -80°C.
El tejido se homogeneizó con un TissueRuptor II antes de extraer el ARN total con el kit RNeasy Plus Mini (74136, QIAGEN), siguiendo los protocolos del fabricante.
Se sintetizó ADNc con el kit High-Capacity RNA-to-cDNA y, posteriormente, se realizó un análisis de la expresión génica con 1-5ng/ml de ADNc, utilizando la mezcla maestra de expresión génica y los ensayos de expresión génica TaqMan, siguiendo las instrucciones de los fabricantes.
Se utilizó el instrumento Applied Biosystems 6500 Fast Real-Time PCR System para procesar las muestras, y la expresión génica relativa se calculó mediante el método 2-ΔCT.
Histología hepática
Las muestras de hígado se fijaron con paraformaldehído al 4% en PBS y se incluyeron en parafina. Tras la desparafinización con xileno y la hidratación mediante un paso por una serie de alcoholes, se tiñeron secciones de 3μm de espesor con hematoxilina-eosina y ácido peryódico de Schiff, antes y después de la digestión con diastasa, en IPATIMUP Diagnostics, Portugal.
La puntuación histopatológica de la actividad necroinflamatoria lobulillar, se asignó utilizando los criterios de Scheuer para la clasificación de la hepatitis crónica. Así, las puntuaciones se asignaron de la siguiente manera: 0 = ausencia de inflamación, 1 = inflamación, pero sin muerte hepatocelular, 2 = necrosis focal (uno o pocos hepatocitos necróticos/cuerpos acidófilos), 3 = muerte focal grave, necrosis confluente sin puentes, y 4 = daño con necrosis en puentes.
Se contabilizó el número total de eventos de citoadherencia endotelial de eritrocitos (secuestración), formación de rosetas y oclusión vascular en campos aleatorios de alta potencia (400 aumentos) de las secciones hepáticas. Las imágenes se capturaron con un fotomicroscopio Olympus BX50.
Para la detección inmunohistoquímica de células CD45+, las secciones hepáticas se sometieron a recuperación de antígeno con tampón de citrato, se bloquearon las peroxidasas endógenas con H₂O₂ al 0,6 % y los antígenos inespecíficos con albúmina sérica bovina al 5 %. Las muestras se incubaron con anticuerpo de cabra anti-ratón CD45, seguido de IgG de conejo anti-cabra conjugada con peroxidasa de rábano picante.
La inmunorreactividad se visualizó mediante 3,3’-diaminobencidina. La cuantificación se realizó contando las células positivas en 5 campos aleatorios por hígado a 200× de aumento utilizando el software QuPath Open para análisis de bioimágenes (versión 0.4.0).

Ensayo de sustancias reactivas al ácido tiobarbatúrico
La peroxidación lipídica y de especies reactivas de oxígeno (ROS) hepáticas, se evaluó cuantificando el malondialdehído mediante el kit de ensayo TBARS, según las instrucciones del fabricante.
Se prepararon homogeneizados de tejido a partir de tejido hepático congelado rápidamente, añadiendo 1ml de tampón RIPA por cada 100mg de tejido y lisando con tubos de homogeneización de tejido blando Precellys, siguiendo las instrucciones del fabricante.
Los lisados se dejaron reaccionar con ácido tiobarbatúrico a 95°C durante 1 hora, se enfriaron en hielo y se centrifugaron durante 10 minutos a 1600 x g a 4°C. Posteriormente, se midió la absorbancia de los lisados a 530nm.
Software y análisis estadístico
Todos los datos de citometría de flujo se analizaron con el software FlowJo (BD). Los gráficos se generaron con GraphPad Prism. Los esquemas de la configuración experimental se crearon con Adobe Illustrator.
El análisis estadístico también se realizó con GraphPad Prism. Se utilizó la prueba de normalidad ómnibus de D’Agostino-Pearson para determinar la normalidad/lognormalidad. Se utilizaron pruebas estadísticas paramétricas para datos con distribución normal. Para distribuciones lognormales, los datos se transformaron mediante logaritmo antes del análisis estadístico. Cuando los datos no presentaban una distribución normal o lognormal, o cuando había muy pocos puntos de datos disponibles para la prueba de normalidad, se aplicó una prueba no paramétrica. Se utilizó una prueba t (o una prueba no paramétrica comparable) para comparar las medias de dos grupos. Se utilizó un ANOVA bidireccional para el análisis con dos variables categóricas y una variable continua.

Resultados del estudio del hierro en las células del hospedador con malaria
En los siguientes apartados, se muestran los resultados obtenidos por los científicos ene le estudio del papel del hierro en las células del hospedador de malaria.
La disminución de la captación de hierro en las células del hospedador aumenta la carga patógena de P. chabaudi.
Para investigar los efectos de la disponibilidad de hierro en las células del hospedador en la respuesta a la malaria, utilizaron un ratón transgénico con una mutación en el transportador celular de hierro TfR1, donde vieron que la mutación TfrcY20H/Y20H disminuye la internalización del receptor en aproximadamente un 50%, lo que resulta en una menor captación celular de hierro.
No obstante, los efectos de la mutación TfrcY20H/Y20H en las células eritroides se minimizan debido a un mecanismo compensatorio mediado por STEAP3. En condiciones de homeostasis, los ratones adultos TfrcY20H/Y20H son sanos, de tamaño normal y no presentan anemia. Sin embargo, tienen eritrocitos microcíticos, lo cual se compensa con un aumento en el número de eritrocitos, y niveles de hierro hepático y sérico ligeramente reducidos.
Se infectaron ratones TfrcY20H/Y20H y de tipo salvaje con una cepa de malaria de roedores transmitida recientemente por mosquitos, P. chabaudi chabaudi AS, que expresa constitutivamente GFP. Se utilizaron parásitos transmitidos recientemente por mosquitos para imitar una infección natural con mayor precisión, ya que se sabe que la transmisión vectorial regula la virulencia de Plasmodium y altera la respuesta inmunitaria del hospedador. En consecuencia, se esperaba que la parasitemia fuera significativamente menor tras la infección con parásitos transmitidos recientemente por mosquitos, en comparación con la infección con parásitos que han pasado por la sangre en serie y que son más virulentos.
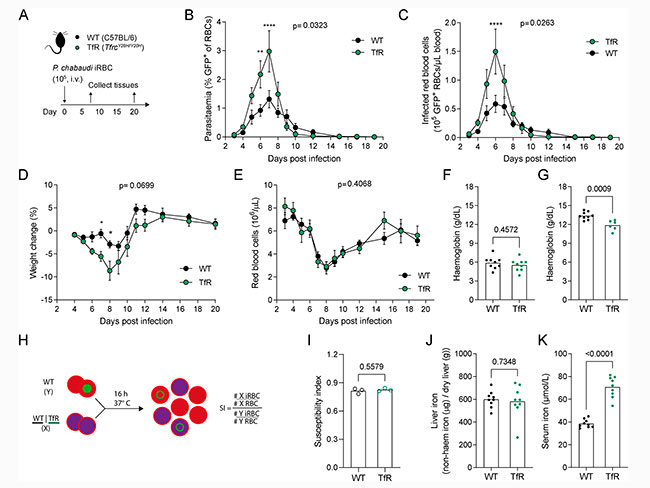
Sorprendentemente, los ratones con una menor captación celular de hierro presentaron una parasitemia máxima significativamente mayor y un mayor número máximo de eritrocitos infectados (iRBC), tal y como se muestra en las Figuras 1B y 1C de la imagen inferior. Además, la mayor carga patógena coincidió con una pérdida de peso más severa que en los ratones de tipo silvestre (Figura 1D).
Este fenotipo contrasta con estudios previos, en los que la deficiencia nutricional de hierro resultó en una menor parasitemia y una mayor supervivencia de los ratones infectados con malaria. Por lo tanto, estos hallazgos resaltan un papel distintivo del hierro en las células en la patología de la malaria, que actúa de forma inversa al efecto protector de la anemia.
Los ratones TfrcY20H/Y20H y los de tipo silvestre presentan una pérdida similar de glóbulos rojos y anemia inducidas por la malaria
Las alteraciones de la fisiología de los glóbulos rojos asociadas a la anemia pueden afectar la infección por malaria y se han propuesto como la causa principal tanto del efecto protector de la deficiencia de hierro como del mayor riesgo asociado a la suplementación con hierro. Por lo tanto, los científicos monitorizaron los glóbulos rojos en ratones de tipo silvestre y TfrcY20H/Y20H infectados con P. chabaudi.
Ambos genotipos mostraron niveles similares de pérdida y recuperación de glóbulos rojos inducidas por la malaria, tal y como se muestra en la Figura 1E de la imagen superior. Además, los ratones TfrcY20H/Y20H y los de tipo silvestre presentaban una anemia igualmente grave en el punto más bajo de pérdida de glóbulos rojos, ocho días después de la infección (dpi) (Figura 1F). En la fase crónica de la infección (20 dpi), los ratones de tipo silvestre mostraron una mejor recuperación de la anemia en comparación con los ratones TfrcY20H/Y20H (Figura 1G), lo que concuerda con una menor capacidad de las células TfrcY20H/Y20H para incorporar hierro.
Si bien la anemia y el recuento de glóbulos rojos fueron comparables entre ambos genotipos durante la infección, era posible que las diferencias en la fisiología de los glóbulos rojos pudieran alterar el curso de la infección. Por consiguiente, realizaron un ensayo de invasión in vitro para determinar si los glóbulos rojos TfrcY20H/Y20H eran más susceptibles a la invasión por P. chabaudi.
Para ello, incubaron in vitro glóbulos rojos de tipo silvestre o TfrcY20H/Y20H marcados con fluorescencia con glóbulos rojos de un ratón de tipo silvestre infectado con P. chabaudi. Tras completar un ciclo de replicación asexual, evaluaron la invasión y calcularon el índice de susceptibilidad (Figura 1H). Los índices de susceptibilidad de los glóbulos rojos de ambos genotipos fueron similares (Figura 1I), lo que indica que la mayor carga parasitaria en los ratones TfrcY20H/Y20H no se debió a una mayor susceptibilidad de sus glóbulos rojos a la invasión de P. chabaudi.
El hierro elevado no altera sustancialmente la infección por P. chabaudi
Además de la anemia, se ha sugerido que las variaciones en los niveles de hierro del hospedador podrían afectar el crecimiento del parásito en la fase sanguínea. Por consiguiente, midieron el hierro hepático no hemo y el hierro sérico en ratones de tipo silvestre y TfrcY20H/Y20H tras la infección por P. chabaudi.
En el pico de la infección, ambos genotipos presentaron niveles elevados de hierro hepático y sérico en comparación con la homeostasis. Los ratones de tipo silvestre y TfrcY20H/Y20H infectados presentaron niveles equivalentes de hierro hepático, pero los niveles de hierro sérico fueron más altos en los ratones TfrcY20H/Y20H (Figuras 1J y 1K).
El elevado hierro sérico observado en los ratones TfrcY20H/Y20H infectados, fue consistente con su capacidad limitada para captar el hierro unido a la transferrina circulante en los tejidos. Sin embargo, decidieron investigar si este hierro sérico suprafisiológico o hiperferremia podría alterar el crecimiento del parásito P. chabaudi.
Para ello, trataron ratones de tipo salvaje con un anticuerpo IgG monoclonal recombinante anti-BMP6 (αBMP6) o un control isotípico. El tratamiento con αBMP6 suprime la expresión de hepcidina y eleva el hierro sérico, como consecuencia de la liberación no regulada de hierro de los depósitos celulares.
Tras esto, los ratones infectados con P. chabaudi tratados con αBMP6 tuvieron un hierro sérico más alto que los ratones tratados con el control isotípico en los días 9 y 21 después de la infección. No obstante, los ratones tratados con αBMP6 y el isotipo tuvieron picos de parasitemia y recuentos máximos de iRBC comparables, aunque los ratones tratados con αBMP6 parecieron eliminar los parásitos de forma ligeramente más eficiente. Además, el tratamiento con αBMP6 no alteró significativamente la pérdida de peso.
En conjunto, estos datos indican que la hiperferremia observada en ratones TfrcY20H/Y20H infectados, no aumenta la parasitemia máxima. Por consiguiente, estos hallazgos sugieren que la captación de hierro en las células no eritropoyéticas es decisiva en la respuesta del hospedador a la malaria.
La disminución de la captación de hierro en las células del hospedador atenúa la respuesta inmunitaria a P. chabaudi.
La respuesta inmunitaria a la malaria controla la parasitemia, y el bazo es el principal órgano de respuesta inmunitaria a la malaria en fase sanguínea. Por lo tanto, evaluaron la respuesta inmunitaria esplénica a P. chabaudi durante la fase aguda de la infección (8 días después de la infección). Curiosamente, los ratones TfrcY20H/Y20H presentaron una esplenomegalia atenuada durante la infección aguda por P. chabaudi, como se muestra en las Figuras 2A y 2B de la siguiente imagen, lo que sugiere una alteración en la respuesta esplénica.
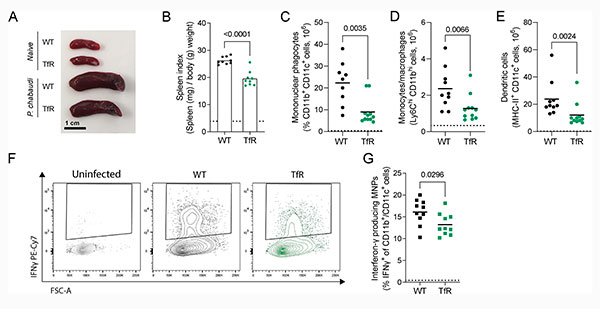
La infección por malaria provoca una afluencia de fagocitos mononucleares (MNP) al bazo, donde participan en la producción de citocinas, la presentación de antígenos y la fagocitosis de eritrocitos infectados. Tras la infección por P. chabaudi, se detectó un menor número de MNP en el bazo de ratones TfrcY20H/Y20H, tal y como se muestra en la Figura 2C de la imagen superior. Esto se observó tanto en los MNP CD11b+ Ly6C+ (similares a monocitos inflamatorios y/o macrófagos derivados de monocitos) como en los MNP CD11c+ MHCII+ (similares a células dendríticas) (Figuras 2D y 2E).
En la infección por malaria, algunas MNP pueden producir IFNγ que facilita la activación y polarización de las células T CD4+ vírgenes. En consecuencia, los esplenocitos de ratones infectados se cultivaron ex vivo con un inhibidor del transporte de proteínas y se realizó una tinción intracelular de citocinas. Curiosamente, menos MNP de ratones TfrcY20H/Y20H produjeron IFNγ, en comparación con las MNP de ratones de tipo salvaje (Figuras 2F y 2G). Sin embargo, los ratones infectados de tipo salvaje y TfrcY20H/Y20H tuvieron números comparables de neutrófilos, eosinófilos y células NK esplénicas durante la infección aguda (8 dpi).
Por lo tanto, los ratones con captación celular de hierro disminuida tuvieron una respuesta atenuada de MNP a la infección por P. chabaudi.
La deficiencia de hierro en las células del hospedador afecta la respuesta de las células T CD4+ a P. chabaudi.
Las células T, en particular las células T CD4+, son un componente crítico de la respuesta inmunitaria a la malaria en fase sanguínea, por lo que, evaluaron la respuesta de las células T esplénicas a la infección aguda por P. chabaudi.
El recuento total de células T CD4+ esplénicas fue comparable en ambos genotipos ocho días después de la infección, como se observa en la Figura 3A de la siguiente imagen. Sin embargo, los ratones con una menor captación de hierro celular presentaron una menor proporción de células T CD4+ efectoras (Figura 3B) y, en consecuencia, un menor número total de células T CD4+ efectoras esplénicas que los ratones de tipo silvestre (Figura 3C). Además, la proporción de células T CD44+ y PD1+ CD4+ con experiencia antigénica también se redujo en los ratones TfrcY20H/Y20H (Figuras 3D y 3E), lo que refuerza su menor activación.
También, se observó una menor cantidad de linfocitos T CD4+ en división activa en ratones TfrcY20H/Y20H, según el marcador de proliferación KI-67 (Figura 3F), lo que sugiere una alteración funcional de la respuesta de los linfocitos T CD4+ a P. chabaudi en ratones con una menor captación celular de hierro.
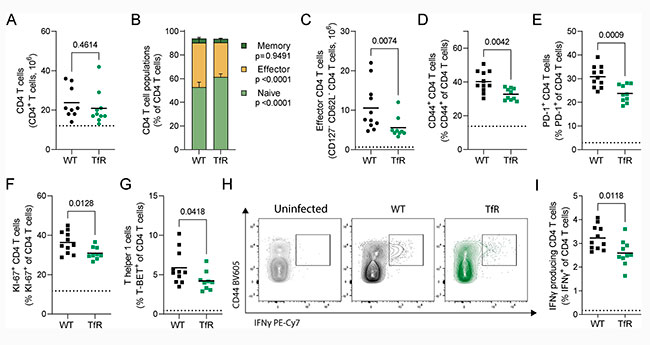
De forma similar, el recuento total de células T CD8+ no difirió entre genotipos, pero los ratones TfrcY20H/Y20H infectados con P. chabaudi presentaron menos células T CD8+ efectoras ocho días después de la infección. Sin embargo, esta vez no se observaron diferencias en el porcentaje de células T CD8+ con experiencia antigénica (CD44+ o PD-1+), proliferantes (KI-67+) o productoras de IFNγ. Por lo tanto, la respuesta de las células T CD8+ a la infección por P. chabaudi también se atenuó, aunque en menor medida que la de las células T CD4+.
Por su parte, las células T helper 1 (Th1) y otros subtipos de células T helper que expresan IFNγ son particularmente importantes para la inmunidad contra la malaria. Curiosamente, la proporción de células T CD4+ que expresaban el factor de transcripción Th1 T-BET fue menor en ratones con una captación celular de hierro reducida (Figura 3G). Además, un menor número de células T CD4+ de ratones TfrcY20H/Y20H produjo IFNγ tras la reestimulación ex vivo (Figuras 3H y 3I). Esto refuerza aún más la evidencia de la alteración funcional de las células T CD4+ en ratones TfrcY20H/Y20H durante la infección por P. chabaudi.
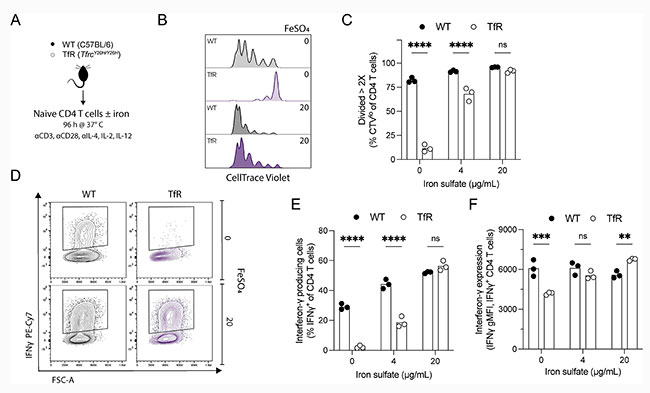
Para determinar si estas deficiencias eran intrínsecas a las células T y dependientes del hierro, utilizaron células T CD4+ vírgenes aisladas de ratones silvestres no infectados y ratones TfrcY20H/Y20H. Las células se cultivaron in vitro en condiciones de polarización Th1 durante cuatro días, en medios de cultivo estándar o suplementados con hierro, como se indica en la Figura 4A de la siguiente imagen.
Los linfocitos TfrcY20H/Y20H pueden adquirir hierro en condiciones donde la transferrina está hipersaturada y es probable que se generen cantidades suficientes de hierro libre. La proliferación se vio significativamente afectada en las células T CD4+ TfrcY20H/Y20H, pero pudo recuperarse de forma dosis-dependiente mediante la suplementación con hierro (Figuras 4B y 4C). Además, muy pocas células T CD4+ TfrcY20H/Y20H cultivadas en medios estándar produjeron IFNγ. Sin embargo, la suplementación con hierro recuperó completamente la producción de IFNγ (Figuras 4D, 4E y 4F). Por lo tanto, las deficiencias de células T CD4+ observadas en ratones TfrcY20H/Y20H durante la infección por P. chabaudi se reprodujeron in vitro y pudieron revertirse mediante la suplementación con hierro.
Estas observaciones confirman que la escasez de hierro en las células del hospedador altera la función de las células T CD4+, lo que conlleva una respuesta reducida de estas células a la infección por P. chabaudi.
La disminución de la captación celular de hierro altera la respuesta del centro germinal a P. chabaudi.
Una respuesta eficiente del centro germinal (CG) es necesaria para generar anticuerpos de alta afinidad que permitan la eliminación de la malaria. En vista de la respuesta deficiente de las células T CD4+ a P. chabaudi en ratones TfrcY20H/Y20H, examinaron con mayor detalle la respuesta de las células T foliculares auxiliares (Tfh) que apoyan a las células B.
Durante la fase aguda de la infección, una menor proporción de células T CD4+ de ratones TfrcY20H/Y20H expresó el receptor de coestimulación de células B ICOS, lo cual se muestra en la Figura 5A de la imagen inferior.
ICOS es esencial en la infección por malaria, ya que es necesario para mantener la respuesta de las células Tfh y la producción de anticuerpos. En consonancia con esto, los ratones TfrcY20H/Y20H presentaron menos células Tfh, tanto durante la fase aguda (8 dpi) como en la crónica (20 dpi) de la infección (Figuras 5B y 5C).
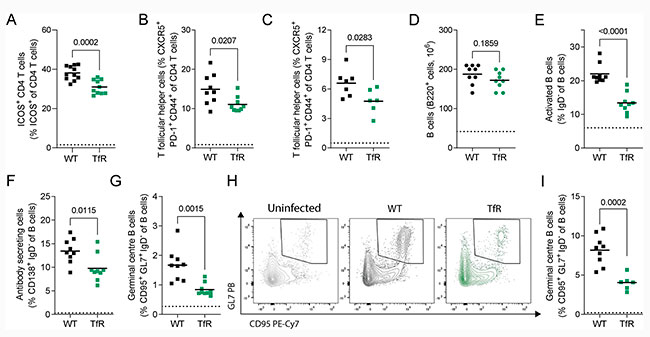
Las células Tfh apoyan la activación, diferenciación y selección de células B del centro germinal de alta afinidad, y son un componente esencial de la respuesta inmune humoral a la malaria. Por lo tanto, a continuación, evaluaron la respuesta de las células B a la infección por P. chabaudi en ratones TfrcY20H/Y20H y de tipo silvestre.
Tras ello, no observaron diferencias entre genotipos en el número total de células B esplénicas en la fase aguda de la infección (8 dpi) (Figura 5D). Sin embargo, los ratones con una captación celular de hierro reducida presentaron una activación de células B gravemente comprometida y menos células B efectoras secretoras de anticuerpos (Figuras 5E y 5F). Además, los ratones TfrcY20H/Y20H tuvieron menos células B del centro germinal durante la infección aguda (8 dpi) (Figura 5G), y este efecto se mantuvo en la fase crónica de la infección (20 dpi) (Figuras 5H y 5I), lo que indica una inhibición inmunitaria prolongada causada por la disponibilidad restringida de hierro en las células del hospedador.
La deficiencia de hierro en las células del hospedador afecta la función de las células B
Para determinar si la mutación TfrcY20H/Y20H también tenía efectos intrínsecos y dependientes del hierro en las células B, investigaron su funcionalidad in vitro. Para ello, aislaron células B de ratones TfrcY20H/Y20H no infectados y de tipo silvestre, y las activaron y cultivaron en medios estándar o suplementados con hierro (citrato férrico amónico (AFeC)) durante tres días, como se indica en la Figura 6A de la siguiente imagen inferior.
Con ello, vieron que la expresión del marcador de activación del transportador de aminoácidos neutros grandes (LAT-1/CD98) en células B divididas (B LAT-1) fue menor en las células B TfrcY20H/Y20H que en las de tipo silvestre. Sin embargo, la suplementación con hierro restauró la expresión de LAT-1, lo que indica una mejor activación de las células B (Figura 6B).
La proliferación de las células B TfrcY20H/Y20H también se vio gravemente afectada en comparación con las células de tipo silvestre, pero se restauró mediante la suplementación con hierro de forma dosis-dependiente (Figuras 6C y 6D).
La escasez de hierro también inhibió el potencial de las células B TfrcY20H/Y20H para diferenciarse en células secretoras de anticuerpos y con cambio de isotipo, y esta alteración se revirtió completamente con la suplementación de hierro (Figuras 6E, 6F y 6G).
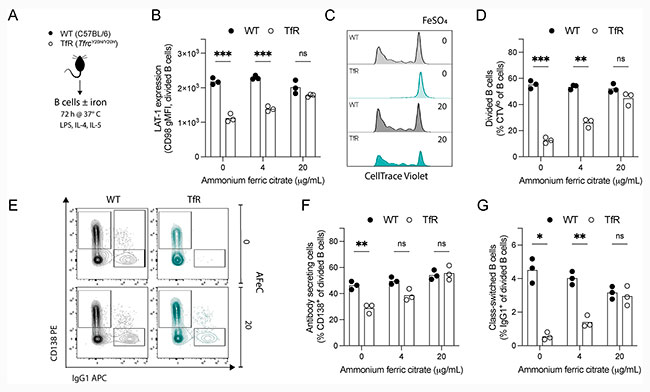
En general, estos datos muestran claramente que la activación, proliferación y diferenciación de las células B TfrcY20H/Y20H se vieron afectadas, lo que demuestra que la deficiencia de hierro celular causa disfunción intrínseca de las células B.
La disminución de la captación celular de hierro atenúa la patología hepática inducida por P. chabaudi
Los ratones TfrcY20H/Y20H presentaron una mayor parasitemia por P. chabaudi y una respuesta inmunitaria inhibida. Sin embargo, las consecuencias precisas de este fenotipo de la enfermedad no estaban claras. Aspectos de la respuesta inmunitaria, como el perfil de citocinas y el equilibrio entre las respuestas proinflamatorias e inmunorreguladoras, pueden inclinar la balanza hacia la protección o la patología en la malaria. Por lo tanto, una respuesta inmunitaria atenuada podría causar hiperparasitemia, pero también podría ser crucial para limitar la inmunopatología. En consecuencia, los autores del trabajo se propusieron caracterizar indicadores clave de la gravedad de la malaria.
En primer lugar, midieron los niveles circulantes de angiopoyetina-2 (ANG-2) y alanina transaminasa (ALT). La ANG-2 es un marcador de activación endotelial que se correlaciona con la gravedad de la malaria y la mortalidad en humanos. Por otro lado, el daño hepático también es indicativo de malaria grave, y la ALT es un marcador estándar de daño hepático.
Tras esto, observaron una tendencia hacia niveles más bajos de ANG-2 y una disminución significativa de la ALT en ratones TfrcY20H/Y20H ocho días después de la infección por P. chabaudi, lo que sugiere una patología más leve, como se muestra en las Figuras 7A y 7B de la imagen inferior.
Dada la diferencia sustancial en la ALT sérica entre los genotipos, examinaron con mayor detalle la patología hepática inducida por la malaria. Así pues, los ratones TfrcY20H/Y20H presentaron una menor expresión de los genes de proteínas de fase aguda Saa1 y Fga, implicados en el daño tisular y la inflamación. Además, si bien ambos genotipos desarrollaron hepatomegalia inducida por la malaria, se observó una tendencia hacia una hepatomegalia menos grave en los ratones TfrcY20H/Y20H.
Por su parte, el análisis histológico reveló patología hepática en todos los ratones infectados con P. chabaudi, caracterizada por necrosis hepatocelular, dilatación sinusoidal, depleción de glucógeno e infiltración de células inmunitarias mononucleares (Figuras 7C y 7D). Curiosamente, no se observó infiltración de células inmunitarias polimorfonucleares.
Todos los ratones silvestres infectados desarrollaron necrosis confluente (áreas de desorganización lobulillar, eosinofilia y pérdida de depósitos de glucógeno), y la mayoría de los individuos (8 de 11) también mostraron necrosis en puente (áreas de necrosis confluente que se extienden a través de múltiples lobulillos) (Figura 7E). En contraste, se detectó necrosis focal grave o necrosis confluente en poco más de la mitad (6 de 10) de los ratones TfrcY20H/Y20H infectados, y solo cuatro individuos desarrollaron necrosis en puente. Por lo tanto, la proporción de ratones que desarrollaron necroinflamación hepática grave tras la infección por P. chabaudi fue significativamente menor en los ratones TfrcY20H/Y20H que en los ratones de tipo silvestre.
Se sabe que el exceso de hierro hepático reactivo y hemo causa daño hepático en la malaria. Sin embargo, no observaron diferencias en el hierro hepático total no hemo ni en la peroxidación lipídica hepática, que se correlaciona con los niveles de especies reactivas de oxígeno. Por consiguiente, es improbable que las variaciones a nivel tisular del hierro hepático reactivo o del hemo puedan explicar la diferencia en el daño hepático.
Además, midieron la expresión de dos genes conocidos por tener un efecto hepatoprotector en el contexto de la sobrecarga de hierro en la malaria: Hmox1 (que codifica la hemoxigenasa-1 (HO-1)) y Fth1 (que codifica la cadena pesada de la ferritina). La expresión génica hepática de Hmox1 fue mayor en ratones TfrcY20H/Y20H, mientras que la expresión de Fth1 no presentó diferencias entre genotipos, ocho días después de la infección. Por lo tanto, la mayor expresión de Hmox1 podría haber contribuido al efecto hepatoprotector en los ratones TfrcY20H/Y20H.
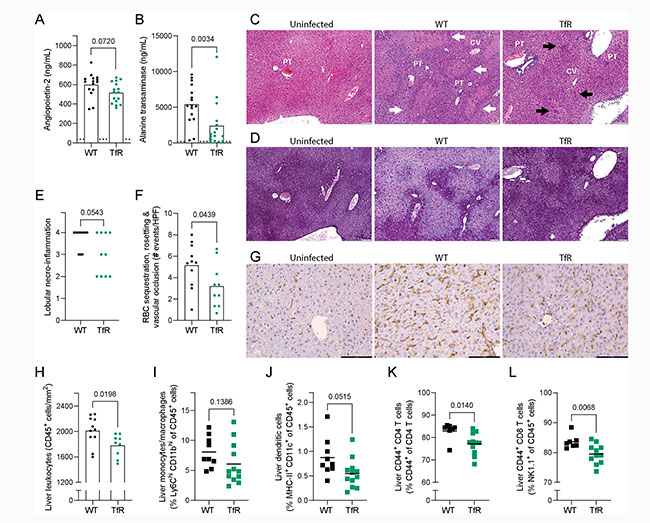
Durante la infección por malaria, la activación endotelial provoca un aumento de la adhesión y el secuestro de eritrocitos infectados, lo que resulta en oclusiones vasculares hepáticas e hipoxia que causan daño. Se detectaron menos eventos de secuestro, formación de rosetas y oclusión vascular en secciones de hígado de ratones TfrcY20H/Y20H ocho días después de la infección por P. chabaudi (Figura 7F). Junto con la tendencia hacia niveles más bajos de ANG-2 en ratones TfrcY20H/Y20H (Fig. 7A), esto indica que la disminución de la activación endotelial y el secuestro de eritrocitos infectados contribuyeron a la atenuación de la patología hepática observada en estos ratones.
La inflamación también causa enfermedad grave y patología hepática en la malaria. Por lo tanto, la inflamación hepática se evaluó midiendo la expresión de genes que codifican las citocinas proinflamatorias IFNγ, TNFα e IL-1β. No observaron diferencias en la expresión de IFNγ o TNFα, pero la expresión de IL-1β fue menor en ratones TfrcY20H/Y20H ocho días después de la infección por P. chabaudi.
Además, la tinción inmunohistoquímica mostró una infiltración reducida de leucocitos (células CD45+) en el hígado de ratones TfrcY20H/Y20H (Figuras 7G y 7H). Asimismo, una menor proporción de leucocitos hepáticos (CD45+) correspondía a células inmunitarias efectoras, como células dendríticas, linfocitos T CD44+ CD4+ y linfocitos T CD8+ CD44+ (Figuras 7I y 7L).
En conjunto, estos datos demuestran que la escasez de hierro en las células del hospedador provoca una respuesta inmunitaria hepática atenuada durante la infección por P. chabaudi.
Discusión y Conclusión
La deficiencia de hierro afecta la infección por malaria en humanos, pero más allá de los efectos de la anemia, se sabía poco sobre cómo la deficiencia de hierro en las células del hospedador influye en la infección por malaria.
Los autores de este estudio, investigaron cómo la adquisición celular restringida de hierro influyó en la infección por P. chabaudi en ratones. Los ratones TfrcY20H/Y20H desarrollaron una anemia inducida por malaria comparable a la de los ratones de tipo silvestre, y la susceptibilidad de los glóbulos rojos a la invasión parasitaria no difirió entre los genotipos. Esto permitió disociar en gran medida los efectos de la anemia de otros efectos del hierro en la respuesta del hospedador a la malaria.
Sorprendentemente, los ratones TfrcY20H/Y20H mostraron una respuesta inmune esplénica y hepática atenuada inducida por P. chabaudi. Esta inhibición inmune se asoció con una mayor parasitemia y una patología hepática atenuada. Por lo tanto, por primera vez, demostraron la función de la adquisición celular de hierro del hospedador a través de TfR1 en la modulación de la respuesta inmune a la malaria, con efectos posteriores tanto en el control del patógeno como en la aptitud del hospedador.
A primera vista, la mayor carga parasitaria observada en ratones TfrcY20H/Y20H podría parecer una consecuencia grave de la deficiencia de hierro en las células del hospedador. Sin embargo, en humanos, una parasitemia elevada no es suficiente para causar una enfermedad grave. Además, el riesgo de malaria grave disminuye significativamente después de solo una o dos exposiciones, mientras que la inmunidad antiparasitaria solo se adquiere después de numerosas exposiciones repetidas. Por lo tanto, mitigar la inmunopatología puede ser más importante que restringir el crecimiento del parásito para la supervivencia del hospedador.
Como señalaron, la mutación TfrcY20H/Y20H tiene consecuencias relativamente leves para los parámetros eritropoyéticos, en comparación con otros linajes hematopoyéticos. Sin embargo, en humanos con captación normal de hierro mediada por TfR1, la deficiencia de hierro suficiente para causar escasez de hierro en las células inmunitarias también suele causar anemia. En tales circunstancias, el crecimiento del parásito probablemente se vería limitado por la anemia, con el resultado final de que la deficiencia de hierro podría ser protectora, en general, si además minimiza aspectos de la inmunopatología.
Trabajos previos han demostrado la importancia de regular los niveles de hemo y hierro en los tejidos para prevenir el daño orgánico en la malaria. Por ejemplo, la HO-1 desempeña un papel importante en la desintoxicación del hemo libre que se produce como resultado de la hemólisis durante la infección por malaria, previniendo así el daño hepático debido a la sobrecarga de hierro tisular, las especies reactivas de oxígeno (ROS) y la inflamación. Curiosamente, los ratones TfrcY20H/Y20H infectados presentaban una mayor expresión de Hmox1, pero niveles de hierro hepático y ROS comparables a los de los ratones de tipo silvestre. En consecuencia, esto podría indicar un mayor procesamiento del hemo que podría tener un efecto protector sobre los tejidos.
En humanos, existe una correlación entre la saturación de transferrina y los niveles de ALT en pacientes con malaria sintomática, lo que sugiere que el estado del hierro podría estar relacionado con la patología hepática inducida por la malaria. Sin embargo, puede resultar difícil interpretar las mediciones del estado del hierro en individuos infectados con malaria, ya que estos parámetros pueden verse alterados por la inflamación y la destrucción de los glóbulos rojos.
Los hallazgos de este trabajo revelan dimensiones adicionales a través de las cuales el estado del hierro del hospedador influye en el daño tisular inducido por la malaria. El daño hepático atenuado que observaron en ratones TfrcY20H/Y20H infectados con P. chabaudi probablemente se explique por una combinación de factores: mayor expresión de HO-1 hepatoprotector, menor activación endotelial mediada por el sistema inmunitario, secuestro de glóbulos rojos infectados y oclusión vascular hepática, así como una inflamación hepática inhibida.
La respuesta inmune proinflamatoria a la malaria tiene efectos posteriores en la citoadherencia, ya que las citocinas proinflamatorias activan las células endoteliales, lo que conduce a una mayor expresión de receptores para la citoadherencia. Como consecuencia, los ratones infectados con P. chabaudi que carecen de inmunidad adaptativa o señalización del receptor de IFNγ, tienen una secuestración sustancialmente reducida de iRBC en el hígado y ningún daño hepático detectable.
Las células endoteliales también pueden activarse por interacciones directas con iRBC, y en humanos, ANG-2 se correlaciona con la biomasa parasitaria estimada. Sin embargo, aunque los ratones TfrcY20H/Y20H infectados con P. chabaudi tuvieron una parasitemia máxima más alta, tuvieron menos secuestración hepática, roseta y eventos de oclusión vascular y niveles más bajos de ANG-2. La atenuación de la respuesta inmunitaria innata y adaptativa es la causa más probable de la disminución de la activación endotelial y la obstrucción microvascular hepática en ratones TfrcY20H/Y20H. Esto, a su vez, probablemente contribuyó a la clara atenuación de la patología hepática, a pesar de la mayor parasitemia.
Tras la infección por P. chabaudi, observaron una extensa infiltración de leucocitos mononucleares en el hígado, pero esta respuesta se vio suprimida en los ratones TfrcY20H/Y20H. Específicamente, los ratones TfrcY20H/Y20H infectados presentaban menos células inmunitarias efectoras en el hígado. Las células inmunitarias hepáticas pueden contribuir al daño hepático en la malaria, por ejemplo, mediante la producción de citocinas proinflamatorias o a través de la destrucción de hepatocitos por efecto indirecto. En consecuencia, una respuesta inmunitaria proinflamatoria hepática más débil probablemente limitó la inmunopatología y atenuó el daño hepático inducido por la malaria en ratones con deficiencia de hierro celular.
En contraste con lo que pasa en la gripe o influenza, la disminución de la adquisición de hierro celular inhibió tanto la respuesta inmunitaria innata como la adaptativa a la malaria, mitigando así el daño del tejido hepático y la inflamación inducidos por la malaria. Esto pone de relieve los complejos efectos de la deficiencia de hierro en el sistema inmunitario y subraya la necesidad de considerar su efecto en diferentes enfermedades infecciosas de manera específica para cada patógeno. Así pues, una mejor comprensión de cómo el estado del hierro del hospedador afecta a la inmunidad, podría beneficiar el desarrollo de terapias antimicrobianas mejoradas y aumentar la seguridad de los tratamientos para la deficiencia de hierro.
La respuesta inmunitaria innata inhibida a P. chabaudi en ratones TfrcY20H/Y20H probablemente contribuyó tanto al aumento de la carga patógena como a la disminución de la patología hepática. Los MNP esplénicos son importantes para controlar la parasitemia, pero también son vitales para mantener la homeostasis tisular y prevenir el daño tisular en la malaria. Aunque otras células innatas, como los neutrófilos, las células NK y las células γδT son una parte importante de la respuesta inmune a la malaria, solo la respuesta de los MNP se vio claramente afectada en ratones TfrcY20H/Y20H.
Cabe destacar que se sabe que los neutrófilos son sensibles a la deficiencia de hierro y que afectan tanto la inmunidad como la patología en la malaria. Sin embargo, en el contexto de P. chabaudi recientemente transmitido por mosquitos, parece que los monocitos y macrófagos, en lugar de los granulocitos, pueden ser particularmente importantes para el control del parásito y la homeostasis tisular.
Las células T CD4+ y las células B se vuelven intrínsecamente disfuncionales durante la escasez de hierro, como demostraron in vitro. Sin embargo, es probable que estos efectos intrínsecos se agraven aún más por las interacciones con otras células con deficiencia de hierro in vivo. Por ejemplo, las células T CD4+ apoyan la respuesta de las células B a la malaria, y la respuesta reprimida de las células T CD4+ a P. chabaudi en ratones TfrcY20H/Y20H presumiblemente limitó aún más la respuesta de las células B.
La proliferación es un aspecto de la función de las células inmunitarias que parece ser particularmente sensible a la deficiencia de hierro. Como era de esperar, también observaron el efecto inhibitorio más significativo en las poblaciones de células inmunitarias que se expanden enormemente durante la infección por P. chabaudi. Además, la proliferación suele ser necesaria para la diferenciación de linfocitos y la función efectora, y la diferenciación de las células Tfh y Th1 en la malaria depende de un subconjunto de células T CD4+ precursoras altamente proliferativas.
Las células T de ratones TfrcY20H/Y20H también mostraron una expresión reducida de KI-67, lo que confirma aún más que la proliferación deficiente es un mecanismo crítico de inhibición inmunitaria en condiciones de escasez de hierro celular.
Las células T CD4+ que producen citocinas proinflamatorias también son sensibles a la restricción de hierro, como demostraron para el IFNγ, y previamente para la IL-2 y la IL-17. Curiosamente, la sobrecarga de hierro también puede alterar la producción de citocinas por las células T CD4+, y el exceso de hierro puede tener un efecto inhibidor sobre la producción de IFNγ. Estas observaciones subrayan que el desequilibrio de hierro en cualquiera de sus extremos puede alterar la función de las células inmunitarias.
A pesar de la mayor parasitemia máxima en los ratones TfrcY20H/Y20H, ambos genotipos fueron capaces de eliminar los parásitos de P. chabaudi a una tasa comparable. Por lo tanto, incluso una respuesta inmunitaria humoral debilitada parece ser suficiente para controlar la infección por P. chabaudi. Sin embargo, este estudio no investigó los efectos de la deficiencia de hierro en las células inmunitarias sobre la formación de inmunidad a largo plazo, la cual podría haberse visto más gravemente afectada. La respuesta deficiente de los glucocorticoides, en particular, sugiere que la deficiencia de hierro podría contrarrestar la formación de una memoria inmunitaria eficaz frente a infecciones posteriores de malaria. Por tanto, futuros estudios deberían considerar si la inmunidad inhibida como resultado de la deficiencia de hierro podría contribuir a este fenómeno. Además, la amplia superposición geográfica y epidemiológica entre la deficiencia de hierro y la malaria, hace que este concepto sea particularmente relevante para futuras investigaciones.
Finalmente, queda por determinar la importancia general del hierro en las células del hospedador en la infección por malaria en humanos, en particular dentro del diverso contexto genético tanto de humanos como de parásitos, presente en las regiones endémicas de malaria. Los modelos murinos de malaria son útiles para generar hipótesis, pero estos hallazgos deben confirmarse y desarrollarse mediante estudios en poblaciones humanas.
Así pues, este estudio reveló que la disminución de la adquisición de hierro por parte de las células del hospedador inhibe la respuesta inmunitaria a la malaria y atenúa el daño hepático, a pesar de una mayor carga parasitaria y un grado similar de anemia en ratones.
En conjunto, estos datos resaltan un papel previamente subestimado del hierro celular en el equilibrio entre el control del patógeno y la inmunopatología, y contribuyen a nuestra comprensión de las complejas interacciones entre la deficiencia de hierro y la malaria. Por lo tanto, estos hallazgos tienen importantes implicaciones para estos dos problemas de salud global, generalizados y urgentes.
Fuente de este artículo
Este artículo fue publicado originalmente en la revista “PLOS Pathogens”, bajo la referencia: Wideman SK, Frost JN, Richter FC, Naylor C, Lopes JM, Viveiros N, et al. (2023) Cellular iron governs the host response to malaria. PLoS Pathog 19(10): e1011679. https://doi.org/10.1371/journal.ppat.1011679; estando disponible con acceso libre en la propia web de PLOS Pathogens.