
Se sabe que la diabetes aumenta la susceptibilidad a las infecciones. En este estudio, sus autores investigaron el efecto de los niveles altos de glucosa en la expresión del péptido antimicrobiano psoriasina y su consecuente aumento de la bacteria E. coli en la vejiga urinaria, lo cual ocasiona infección de orina derivada de la diabetes.
Presentación
“Diabetes downregulates the antimicrobial peptide psoriasin and increases E. coli burden in the urinary bladder” (“La diabetes regula negativamente el péptido antimicrobiano psoriasina y aumenta la carga de E. coli en la vejiga urinaria.”), es un artículo científico elaborado por Mohanty, S., Kamolvit, W., Scheffschick, A., Björklund, A., Tovi, J., Espinosa, A., Brismar, K., Nyströn, T., Schröder, J. M., Östenson, C., Aspenström, P., Brauner, H. y Brauner A.; y publicado el 20 de Setiembre de 2022 en el volumen 13 de la revista Nature Communications; el cual trata sobre la infección de orina derivada de la diabetes, en el que sus autores investigaron el efecto de los niveles elevados de glucosa y de diabetes en la expresión del péptido antimicrobiano psoriasina y sus posibles consecuencias para la infección urinaria por E. coli.
La alta prevalencia de diabetes representa un importante desafío para la salud mundial, a menudo acompañada de un mayor riesgo de infecciones bacterianas, en parte debido al deterioro de la función granulocítica y a cambios en la inmunidad innata.
Dentro de las infecciones derivadas de la diabetes, las del tracto urinario (ITU) causadas por E. coli son comunes y se asocian con mayor frecuencia a complicaciones graves, como la urosepticemia. Tradicionalmente, se ha considerado que la glucosuria proporciona un entorno óptimo para el crecimiento bacteriano, sin embargo, diversos factores como la edad, un control metabólico deficiente y complicaciones a largo plazo, contribuyen al riesgo de infección de orina derivada de la diabetes. Además, el deterioro inmunogénico influye en la patogénesis de las ITU graves en pacientes diabéticos.
Por su parte, las células hospedadoras cuentan con mecanismos de defensa que las protegen contra los microorganismos invasores. Recientemente, se han identificado varios factores que pueden proteger la vejiga de patógenos invasores. Los péptidos antimicrobianos (AMP), parte de la respuesta inmunitaria innata de primera línea, defienden el urotelio de los patógenos y tienen el potencial de convertirse en nuevas terapias para las ITU. Se expresan en células epiteliales y endoteliales, así como en células inmunitarias como los neutrófilos. En concreto, el péptido antimicrobiano psoriasina, codificado por S100A7 y perteneciente a la familia de proteínas S100, se ha detectado en el tracto urinario. Este es conocido principalmente por su alta actividad antibacteriana contra E. coli, secuestrando zinc, lo cual restringe el crecimiento bacteriano.
Si bien se reconoce la importancia de la psoriasina durante las infecciones por E. coli, aún se desconoce su posible actividad en la diabetes y en casos de hiperglucemia. Por ello, en este estudio, sus autores investigaron el impacto de la glucosa sobre la psoriasina y la patogénesis de la infección de orina derivada de la diabetes por E. coli, con énfasis en el uroepitelio y las estrategias de defensa en la vejiga urinaria durante la diabetes.
Para ello, estudiaron muestras de sangre, orina y células exfoliadas de orina de pacientes. Examinaron la influencia de la glucosa y la insulina durante el pinzamiento hiperglucémico en individuos con prediabetes y en pacientes con diabetes tipo 1, así como euglucémicos e hiperinsulinémicos, sometidos al pinzamiento. También confirmaron hallazgos importantes in vivo en ratones con diabetes tipo 2 y lo verificaron en líneas celulares uroepiteliales humanas.
Tras esto, vieron que las concentraciones elevadas de glucosa inducen niveles más bajos de psoriasina y deterioran la función de la barrera epitelial, además de alterar las proteínas de la membrana celular y los elementos del citoesqueleto, lo que resulta en un aumento de la carga bacteriana.
El tratamiento con estradiol restaura la función celular al aumentar la psoriasina y la destrucción bacteriana en las células uroepiteliales, lo que confirma su importancia durante la infección del tracto urinario con hiperglucemia.
Métodos
En los siguientes apartados se indican los métodos llevados a cabo por los investigadores para obtener todo el material de trabajo y los datos a partir de su estudio.
Participantes del estudio y recolección de células exfoliadas de suero, plasma y orina humanos
El estudio fue aprobado por el Comité Regional de Ética de Estocolmo y se obtuvo el consentimiento informado de todos los pacientes y voluntarios participantes. Se incluyeron adultos con prediabetes, diabetes mellitus (DM) y controles no diabéticos. Se excluyeron los pacientes y voluntarios con ITU activa, tratamiento antimicrobiano o suplementación con estrógenos. Los individuos con prediabetes no tomaban ningún medicamento antidiabético y presentaban glucemia en ayunas alterada (GAA), intolerancia a la glucosa (IGT) o ambas, GAA e IGT, según una prueba de tolerancia a la glucosa oral. Tanto la GAA como la IGT confieren un alto riesgo de desarrollar diabetes tipo 2.
Se incluyó el plasma de pacientes estudiados durante pinzamientos hiperinsulinémicos euglucémicos y de individuos con prediabetes durante pinzamientos hiperglucémicos.
La glucemia venosa, la HbA1c y la creatinina en orina, se analizaron en el Laboratorio del Hospital Universitario Karolinska siguiendo un protocolo estándar.
Cepa bacteriana
Se utilizó la cepa uropatógena de E. coli CFT073 para experimentos in vitro e in vivo. Esta cepa se aisló de un paciente con pielonefritis aguda.
Las bacterias se cultivaron durante la noche en placas de agar sangre a 37°C, seguidas de 4 horas en caldo Luria-Bertani hasta alcanzar la fase logarítmica. Se lavaron dos veces con solución salina tamponada con fosfato (PBS), y se midió la concentración bacteriana espectrofotométricamente, confirmándose mediante recuento de bacterias viables.
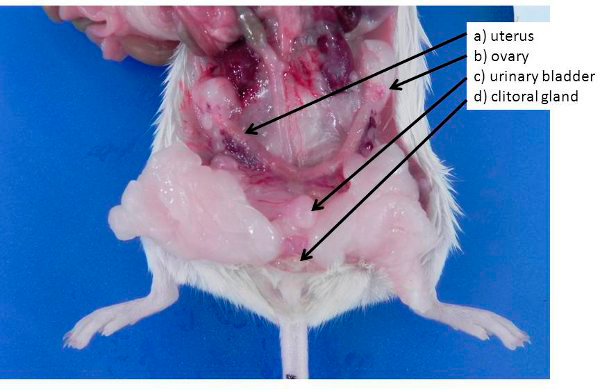
Modelo de ratón de infección urinaria
Los experimentos con ratones fueron aprobados por el Comité de Ética Animal del Norte de Estocolmo y se llevaron a cabo de acuerdo con las directrices de la Federación de Asociaciones de Ciencias de Animales de Laboratorio y en cumplimiento de los requisitos del Comité.
Se obtuvieron ratones hembra db/db (BKS (D)-Leprdb/JOrlRj) de ocho semanas de edad con diabetes tipo 2 y ratones C57BL/6j de tipo salvaje de Janvier Laboratories. Todos los ratones se mantuvieron en una instalación específica libre de patógenos hasta que en la semana 10 se les realizó la infección.
Los ratones se anestesiaron con isoflurano y se infectaron transuretralmente con 0,5 × 108 unidades formadoras de colonias (UFC) de E. coli CFT073 en 50µl de PBS. Se midieron los niveles de glucosa en sangre antes de la infección, durante el curso de la infección y en el sacrificio después de 24h, 7 o 14 días de infección. Se recolectó orina y las vejigas urinarias respectivas se extrajeron asépticamente, se abrieron y se lavaron con PBS para eliminar la orina y las bacterias no adherentes. Para determinar la carga bacteriana total, las bacterias adheridas e intracelulares, las vejigas se homogeneizaron en 1ml de PBS, se diluyeron en serie y, tanto las vejigas como la orina, se sembraron en placas de agar sangre.
Líneas celulares y condiciones de cultivo
Las células uroepiteliales humanas inmortalizadas con telomerasa, TERT-NHUC y 5637, se cultivaron en medio EpiLife con un 1% de suplemento de crecimiento de queratinocitos humanos, y en medio RPMI 1640 sin glucosa, pero suplementado con 5mM de glucosa (Sigma) y un 10% de suero bovino fetal, respectivamente, a 37°C y un 5% de CO2.
Para simular la hiperglucemia, las células se expusieron a 11 y 30mM de glucosa durante 24 a 72h según fuera apropiado, mientras que la normoglucemia, 6mM, se obtuvo con medios de cultivo para TERT-NHUC, y las células 5637 se suplementaron sin citotoxicidad ni comprometiendo las proliferaciones.
Para los experimentos con estradiol, se utilizó medio libre de rojo fenol y suero bovino fetal tratado con carbón al 5%, así como también se utilizó 17β-estradiol (Sigma) en etanol absoluto a una concentración final de 10nM. Las células se trataron durante 48h, con cambio de medio después de 24h. Y para los experimentos con latrunculina B, las células se trataron con 1µM de Lat B 1h antes de añadir glucosa y estradiol.
La infección se inició después de 24h. Durante el experimento de quelación intracelular de zinc, se añadieron 20µM de N,N,N′,N′-tetrakis (2-piridilmetil) etilendiamina a células uroepiteliales humanas TERT-NHUC, durante 2h en medio esencial mínimo, seguido de un tratamiento con alto contenido de glucosa durante 24h en medio Epilife.
Ensayos de infección celular
Los experimentos celulares se llevaron a cabo en placas de cultivo celular de 24 pocillos, con células uroepiteliales humanas TERT-NHUC en Primaria (BD) y las células 5637 en placas Costar. Las células se cultivaron en presencia de glucosa 5 o 6, 11 y 30mM, respectivamente.
Se añadieron 106 UFC/ml (MOI 5) o 2 × 106 UFC/ml (MOI 10) de E. coli CFT073 a células pretratadas casi confluentes y se incubaron a 37°C con 5% de CO2 y 80% de humedad. A los 5, 15, 30, 60 y 120 minutos, las células se lavaron una vez con PBS y se recolectaron para su posterior análisis.
Ensayo de adhesión y supervivencia
Las células TERT-NHUC y 5637 se infectaron con 106 UFC de E. coli en 100µl de PBS por pocillo y se centrifugaron durante 1 min a 350g.
Para evaluar la adhesión bacteriana, las células se infectaron solo durante 30 min y se lavaron con PBS.
En los ensayos de supervivencia, las células se lavaron con PBS tras 2h de infección para eliminar las bacterias no adherentes y se suplementaron con medio fresco durante otras 2h. En las células hiperglucémicas, el medio se suplementó con 30mM de glucosa durante todo el experimento. En algunos puntos temporales, las células se lisaron con Triton-X-100 al 0,1% en PBS, se diluyeron en serie y se sembraron en placas de agar sangre.
La tasa de adhesión y supervivencia se calculó mediante el número de bacterias adheridas o intracelulares, en relación con el número total del mismo experimento.
Actividad antimicrobiana, efecto de la psoriasina
Las células uroepiteliales humanas TERT-NHUC se trataron con glucosa 6 o 30mM. Tras 24h de tratamiento, se retiró el medio y las células se lisaron en Triton X-100 al 1% en PBS. El sobrenadante acelular se obtuvo mediante centrifugación a 8000 g durante 5 min y, posteriormente, se incubó durante 30 min a 37°C con 1µg/ml de anticuerpo monoclonal de ratón antipsoriasina o la misma concentración de un anticuerpo isotipo de control.
A continuación, se añadieron 50µl de una suspensión bacteriana de 10⁻¹ UFC/ml a 150µl de lisado celular pretratado. Tras 30 minutos de incubación a 37°C, se sembraron alícuotas de 100µl y se determinó la supervivencia bacteriana mediante recuento de células viables. Los resultados se expresaron en relación con los lisados celulares control pretratados con anticuerpos de control.
Se recogieron muestras de orina sin crecimiento bacteriano de controles no diabéticos y pacientes con diabetes tipo 2. Se preparó E. coli CFT073 y se añadieron 50µl de una suspensión bacteriana de 10⁻¹ UFC/ml a 150µl de orina con 5µM de péptido de psoriasina. Después de una incubación de 30 minutos a 37°C, se sembraron alícuotas de 100µl y se determinó la supervivencia bacteriana mediante un recuento de bacterias viables.

Aislamiento de ARN total y PCR en tiempo real
Tras la incubación, las células uroepiteliales humanas se procesaron directamente para el aislamiento de ARN, mientras que los tejidos vesicales se homogeneizaron manualmente.
La concentración y pureza del ARN se determinó con nanodrop, y hasta 0,5μg de ARN se transcribieron de forma inversa. Posteriormente, se transcribieron a ADNc en un termociclador. Se realizó PCR en tiempo real después de la desnaturalización inicial a 95°C durante 10 minutos.
Inmunofluorescencia de secciones y células de vejiga de ratón
En el momento requerido, el tejido vesical se fijó en 1ml de PFA al 4% durante al menos 36h y, a continuación, se transfirió a 1ml de etanol absoluto.
Se prepararon bloques de parafina y se cortaron secciones de 4µm con un micrótomo. Las secciones de tejido vesical de ratón incluidas en parafina se desparafinaron y rehidrataron.
Las células uroepiteliales humanas TERT-NHUC se fijaron en PFA al 4% durante 15 minutos a temperatura ambiente y se permeabilizaron con Triton X-100 al 0,3% en PBS. Posteriormente, las secciones se bloquearon durante 30 minutos con FX Signal Enhancer, y tanto las células como las secciones se bloquearon durante 60 minutos adicionales con los sueros de las especies en las que se generaron los anticuerpos secundarios.
La incubación con anticuerpos primarios se realizó durante la noche a 4°C, seguida de anticuerpos secundarios conjugados con Alexa Fluor durante 1 hora a temperatura ambiente.
Las secciones y las células se montaron con medio de montaje que incluía DAPI. Para la estimación nuclear de YAP/TAZ, se utilizó el módulo de densidad integrado en ImageJ para medir la intensidad de fluorescencia total de cada célula, seguida de la intensidad de fluorescencia en el núcleo de cada célula. Se calculó la proporción nuclear sobre el total celular YAP/TAZ.
El análisis del tipo de organización del filamento de actina se realizó analizando imágenes adquiridas por microscopía de inmunofluorescencia, para detectar la presencia de haces cortos de actina, fibras de estrés o actina cortical como forma dominante de organización de la actina. La cuantificación se realizó a partir de tres experimentos independientes por condición experimental; y la intensidad de fluorescencia por unidad de área se analizó con el software Image J Fiji 1.53b.
Prueba ELISA de psoriasina, ocludina e IL-6
Se recogieron muestras de suero y plasma de pacientes con diabetes, junto con muestras de control prediabéticas con pinzamiento hiperglucémico y de pacientes con diabetes tipo 1 con pinzamiento hiperinsulinémico, y muestras de control no diabéticas, antes y después de 2h de tratamiento con glucosa o insulina, respectivamente.
Se recogieron los sobrenadantes de células TERT-NHUC tratadas con glucosa durante 24h, se centrifugaron a 350 g durante 10 min y se almacenaron a -80°C hasta su análisis.
Se utilizaron células no infectadas, suero y plasma obtenidos de individuos no diabéticos antes de la infusión de glucosa e insulina como controles.
Las pruebas ELISA se analizaron con el kit CircuLex S100A7/Psoriasin ELISA (MBL International) e IL-6 (R&D Biosystems), en el lector de microplacas EZ400 y el software ADAP versión 2.0.
Citometría de flujo
Para investigar la expresión proteica de psoriasina, ocludina y caveolina 1, se recogieron células TERT-NHUC tras 36h de tratamiento con glucosa.
Para MRC1 (CD206), las células se infectaron durante 2h y se centrifugaron a 350 g durante 3 min a temperatura ambiente (TA). Se añadió 1ml de PFA al 4% en PBS al sedimento celular. Las células se incubaron a TA durante 15 min, se centrifugaron y se añadió 1ml de Triton-X-100 al 0,1% en PBS (PBST) y se incubaron a TA durante 10 min. Posteriormente, las células se bloquearon con BSA al 5% durante 30 min y se tiñeron con el anticuerpo primario en una proporción 1:1 de 200µl de PBST 1× y BSA al 5% durante 30 min a temperatura ambiente. Tras esto, las células se lavaron con PBS 1× con BSA al 1% y se incubaron con los respectivos anticuerpos secundarios Alexa fluor 488 o Alexa fluor 647, durante 25 min en oscuridad a temperatura ambiente. Finalmente, las células se disolvieron en PBS y los datos se adquirieron en un BD LSRFortessa™ y se analizaron con el software FlowJo versión 10.8.1.

Deleción de S100A7 mediante el sistema crispr/cas9
Se cultivaron células uroepiteliales humanas TERT-NHUC en placas de 24 pocillos hasta una confluencia del 50-70% y se transfectaron con SpCas9/gRNA no dirigido o con un grupo de SpCas9/gRNA específico para S100A7, utilizando el reactivo de transfección Cas9 Lipofectamine™ CRISPRMAX™ (ThermoScientific), siguiendo las instrucciones del fabricante.
Tras la transfección, las células TERT-NHUC se cultivaron durante 72h y después se fijaron y procesaron para el análisis microscópico de anti psoriasina de ratón y anti ocludina de conejo.
Transfección de RHOA/B
Se cultivaron células uroepiteliales humanas 5637 en una placa de 24 pocillos. Al alcanzar una confluencia del 70%, las células se pretrataron con alto contenido de glucosa durante al menos 6h antes de la transfección.
Se preparó por separado 1μg de RHOA/T19N y RHOB/T19N marcados con Myc en 150mM de NaCl hasta un volumen final de 50μl. Se añadieron 2μl de reactivo jetPEI® en 150mM de NaCl hasta un volumen final de 50μl. Se añadieron 50μl de solución jetPEI® de una sola vez a los 50μl de solución de ADN y se incubaron durante 20 min a temperatura ambiente.
Se añadieron gota a gota 100μL de la mezcla jetPEI®/ADN a las células en 1ml de medio con suero y se homogeneizaron mediante agitación suave de la placa. Las células se incubaron durante 24h, seguidas de su fijación con 1ml de PFA al 4% y se procesaron para el análisis microscópico de anti-YAP/TAZ de ratón, detectado mediante anticuerpo de conejo anti-ratón conjugado con Alexa Fluor 488, y de RHOA/T19N y RHOB/T19N marcados con Myc, detectados mediante anticuerpo de conejo anti-Myc, seguido de anticuerpo de burro anti-conejo conjugado con Alexa Fluor 350, utilizando el protocolo estándar de imagenología.
Péptidos
Se utilizaron péptidos de psoriasina 15 de 11.366 Da, libres de Zn2+ purificados y derivados de piel natural, e IL-1β humana recombinante e IL-6, para estimular TERT-NHUC durante 24h.
El bloqueo de IL-1β se logró mediante la adición de diacereína, un inhibidor específico de IL-1β, durante al menos 4h antes del tratamiento con péptidos de IL-1β, seguido de un tratamiento nocturno en TERT-NHUC tratado con niveles altos de glucosa.
Análisis estadístico
Todas las pruebas estadísticas se realizaron en Graphpad Prism versión 5. No se excluyeron muestras de estudios en humanos ni en animales. Para el análisis in vitro con células uroepiteliales humanas, se excluyeron los valores atípicos definidos mediante la prueba de Grubb.

Resultados
A continuación, se comentan los resultados que obtuvieron los investigadores tras el análisis de los datos de este estudio sobre infección de orina derivada de la diabetes.
La glucosa alta disminuye la psoriasina en suero/plasma y vejiga urinaria
Se analizó el posible impacto de la glucosa alta en los péptidos antimicrobianos en células uroepiteliales humanas TERT-NHUC, lo que resultó en una expresión significativamente menor de psoriasina (S100A7) en comparación con la glucosa baja.
Para garantizar la relevancia clínica, investigaron los niveles de psoriasina en pacientes con diabetes, en los que observaron una regulación negativa del ARNm de S100A7 en las células exfoliadas en orina, así como niveles más bajos de proteína psoriasina en orina, con una tendencia similar en los niveles séricos de psoriasina, en comparación con los controles no diabéticos, lo cual se muestra en la siguiente Figura 1a y 1b.
Para investigar el posible impacto de la glucosa y la insulina en la psoriasina, se utilizaron pinzas hiperinsulinémicas hiperglucémicas y euglucémicas. Las pinzas hiperglucémicas se realizaron en individuos prediabéticos con un alto riesgo de desarrollar diabetes tipo 2 (DT2). En estos estudios de pinzamiento, la infusión de glucosa durante 2h resultó en un aumento de la glucosa plasmática media de 5,4mM a 11,4mM. Esto estimuló la secreción de insulina, aumentando los niveles plasmáticos medios de insulina de 16,1mU/L a 95,2mU/L. Y estos cambios se acompañaron de una disminución de los niveles de psoriasina (Figura 1c).
Por el contrario, las pinzas hiperinsulinémicas euglucémicas en pacientes con diabetes tipo 1 (DT1) y en individuos no diabéticos, no mostraron diferencias en los niveles plasmáticos de psoriasina (Figura 1d). Por lo tanto, los niveles plasmáticos más bajos de psoriasina se asociaron con niveles elevados de glucosa en sangre, pero no con niveles de insulina.
Para confirmar los resultados in vivo, se estudiaron ratones hembra db/db con DT2 y ratones control C57BL/6j no diabéticos. Los niveles de ARNm de S100a7a (Figura 1e) y de proteína psoriasina (Figura 1f) fueron significativamente menores en las vejigas de ratones diabéticos, localizándose el péptido principalmente en las capas superficiales de células paraguas.
Para investigar más a fondo el efecto sobre la psoriasina durante la infección en la diabetes, se infectaron transuretralmente ratones con E. coli y observaron una menor expresión de S100a7a en el ARNm (Figura 1g) y en los niveles de proteína (Figura 1h) a las 24h y a los 7 días postinfección en vejigas urinarias de ratones diabéticos, en comparación con los ratones control.
De igual manera, las células uroepiteliales TERT-NHUC cultivadas en condiciones normales de glucosa mostraron un rápido aumento del pico de ARNm de S100A7 ya después de 15 min de infección por E. coli (Figura 1i). Por otro lado, las células TERT-NHUC cultivadas en condiciones de glucosa elevada expresaron un menor nivel de ARNm de S100A7, con resultados similares en células 5637 uroepiteliales. De acuerdo con datos de ARNm, también observaron niveles más bajos de psoriasina asociada al citoplasma y a las vesículas, así como proteína psoriasina secretada (Figura 1k) en las células TERT-NHUC tratadas con alto contenido de glucosa.

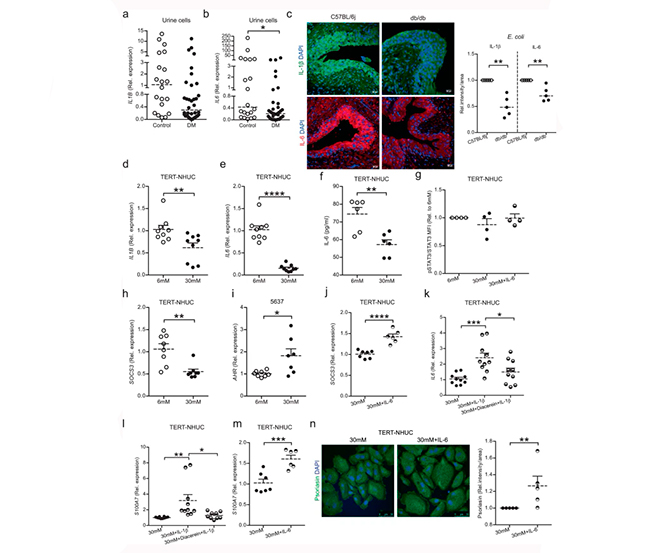
La glucosa alta altera la expresión de psoriasina mediada por IL-6
Dado que se sabe que las citocinas regulan los péptidos antimicrobianos, plantearon la hipótesis de que no solo la psoriasina, sino también las citocinas, podrían verse afectadas por la glucosa alta. En consonancia con esto, las citocinas proinflamatorias IL1B (Figura 2a) e IL6 (Figura 2b) mostraron una regulación negativa del ARNm en células exfoliadas de orina de pacientes con diabetes. Además, los niveles proteicos de IL-1β e IL-6 fueron más bajos y se localizaron en las células paraguas superficiales (Figura 2c) en vejigas de ratones diabéticos 24h después de la infección por E. coli. Asimismo, en células uroepiteliales TERT-NHUC tratadas con glucosa alta, la expresión del ARNm de IL1B e IL6 (Figuras 2d, e) y de la proteína IL-6 (Figura 2f) se vio comprometida.
Aunque IL-6 es un regulador conocido de pSTAT3/STAT3, no se observaron diferencias en pSTAT3 entre células TERT-NHUC tratadas con niveles altos y bajos de glucosa, sustituidas con 50ng/ml de péptido IL-6 humano (Figura 2g).
En las células tratadas con niveles altos de glucosa, la expresión del ARNm de SOCS3, dependiente de STAT3, se vio afectada (Figura 2h), posiblemente debido al aumento de la expresión del ARNm del receptor de hidrocarburos arílicos (AHR) en las células 5637 (Figura 2i), que se sabe que regula negativamente SOCS3 y también aumenta con niveles altos de glucosa. Curiosamente, el ARNm de SOCS3 se incrementó mediante el tratamiento con péptido IL-6 (Figura 2j) en células TERT-NHUC. Para confirmar aún más el efecto de IL-1β en la expresión de IL-6, la suplementación con 20ng/ml de péptido IL-1β resultó en un aumento de la expresión de ambos ARNm de IL6 y S100A7, que fue inhibido por diacereína, un bloqueador específico de IL-1β (Figuras 2k, l).
Se ha demostrado que las expresiones de IL-6 y psoriasina dependen una de la otra en varias células; por lo tanto, especularon que la glucosa alta conduciría a niveles más bajos de IL-6 que podrían afectar la expresión de psoriasina. Por lo tanto, las células TERT-NHUC tratadas con glucosa alta se trataron con péptido IL-6 recombinante y demostraron así que la psoriasina se reguló positivamente, tanto en los niveles de ARNm (Figura 2m) como de proteína (Figura 2n), lo que confirma la interrelación entre IL-6 y psoriasina también durante la hiperglucemia.
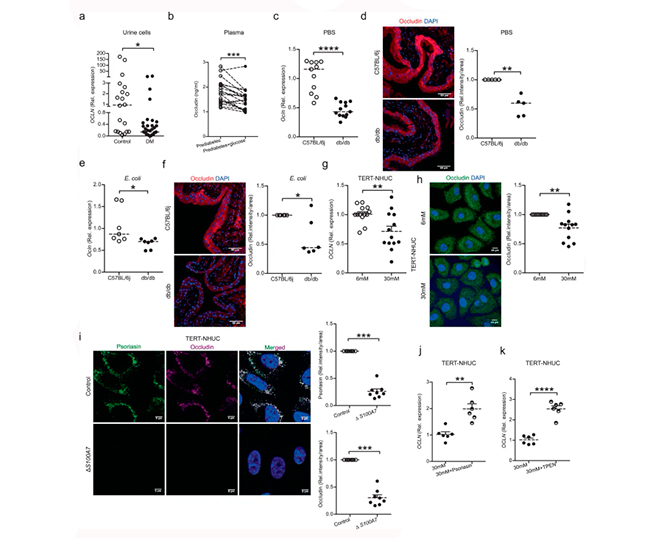
Los niveles altos de glucosa comprometen la expresión de ocludina en plasma y vejiga
Se ha demostrado que los péptidos antimicrobianos regulan la función de la barrera epitelial. Por lo tanto, se planteó que los niveles más bajos de psoriasina podrían afectar la expresión de la proteína de unión estrecha, ocludina. En consonancia con esta hipótesis, las células exfoliadas en orina de pacientes con diabetes, que mostraron niveles más bajos de psoriasina, presentaron una clara disminución de la expresión de OCLN. Asimismo, el plasma de individuos con prediabetes e hiperglucemia sometidos a pinzamiento, mostró niveles reducidos de ocludina, lo cual se muestra en las siguientes Figuras 3 a y b.
De manera similar, observaron un nivel más bajo de ocludina en niveles de ARNm (Figura 3c) y proteína en los ratones diabéticos vs. no diabéticos, con la proteína localizada en las capas superficiales superiores de células paraguas de la vejiga (Figura 3d), a las 24h (Figuras 3e, f) y a los 7 días después de la infección por E. coli. Además, estos resultados se confirmaron in vitro, donde las células uroepiteliales, TERT-NHUC, expuestas a alta glucosa regularon negativamente la ocludina (Figura 3g) a nivel de ARNm y proteína (Figura 3h).
Para confirmar el papel de la psoriasina en la expresión de ocludina, se eliminó S100A7 en células TERT-NHUC utilizando el sistema crispr/cas9, resultando en una menor expresión de ocludina (Figura 3i) sin inducir ningún efecto adverso en las células, evidentemente aparte de la integridad nuclear y del citoesqueleto.
Además, para verificar el efecto de la psoriasina, las células uroepiteliales TERT-NHUC tratadas con glucosa alta y suplementadas con péptido de psoriasina adicional, mostraron una mayor expresión del ARNm de OCLN (Figura 3j).
La forma reducida de Cys de la psoriasina es un potente quelante de zinc endógeno. Por lo tanto, especularon que la psoriasina podría tener una función reguladora en la homeostasis del zinc y que sus propiedades de unión al zinc afectarían la expresión del gen de la ocludina. Para probar si la propiedad de unión al zinc de la psoriasina podría explicar los efectos observados, se utilizó el quelante de penetración celular y específico para el zinc, N,N,N’,N’-tetrakis (2-piridilmetil) etilendiamina (TPEN). Así pues, las células uroepiteliales humanas, TERT-NHUC, se trataron con TPEN, seguido de glucosa alta durante un total de 24h. Esto resultó en un aumento del ARNm de OCLN (Figura 3k) similar al efecto del péptido psoriasina (Figura 3j), lo que indica un posible papel de la disminución intracelular de zinc en el aumento de la expresión de OCLN en células uroepiteliales humanas tratadas con alto contenido de glucosa.
La glucosa alta y la infección de orina derivada de la diabetes por E. coli modulan las proteínas de membrana
Dado que las células TERT-NHUC expuestas a niveles altos de glucosa no mostraron una respuesta pronunciada a la psoriasina al ser infectadas con E. coli, se especuló qué otros factores involucrados en la infección bacteriana también podrían verse afectados.
En este sentido, se sabe que los receptores de manosa o similares a la manosa (MRC1) desempeñan un papel importante en la adhesión bacteriana a las células uroepiteliales. En el estado no infectado, la expresión de Mrc1 fue similar en las vejigas de ambos tipos de ratones. Sin embargo, después de 24h de infección por E. coli, los ratones control diabéticos, pero no los no diabéticos, mostraron una expresión significativamente mayor de Mrc1, tanto en el ARNm como en proteína. En consonancia con estos resultados, las células uroepiteliales TERT-NHUC tratadas con glucosa alta e infectadas con E. coli mostraron una mayor expresión de MRC1 en el ARNm y en proteína, lo cual se muestra en la siguiente Figura 4 a y b.
Tras la unión bacteriana a las células uroepiteliales, la caveolina 1 influye en la captación endocítica de E. coli mediante la formación de caveolas en forma de matraz. En consecuencia, las vejigas de ratones diabéticos mostraron una clara regulación positiva de la caveolina 1 (Figura 4c), con un resultado similar en las células uroepiteliales TERT-NHUC tratadas con altos niveles de glucosa (Figura 4d).
Con base a estos hallazgos, plantearon la hipótesis de que no solo la glucosa, sino posiblemente también la psoriasina, podría afectar a las proteínas de membrana. Sin embargo, el péptido de psoriasina no tuvo efecto sobre MRC1 en las células TERT-NHUC, mientras que la sustitución revirtió la regulación positiva inducida por la hiperglucemia de CAV1 (Figura 4e), lo que sugiere que al añadir psoriasina el efecto de los altos niveles de glucosa podría verse superado.
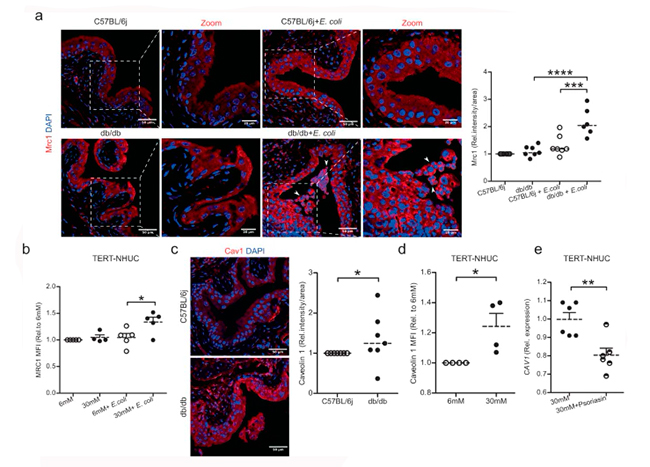
Los niveles altos de glucosa reorganizan el citoesqueleto y translocan YAP/TAZ al núcleo
Se sabe que las Rho GTPasas y la caveolina 1 median los reordenamientos del citoesqueleto de actina. Las proteínas Rho pueden regularse localizándose en las caveolas e interactuando con las caveolinas. Dado el efecto sobre la caveolina 1, especularon que los niveles altos de glucosa y la psoriasina también podrían influir en RhoB y el citoesqueleto.
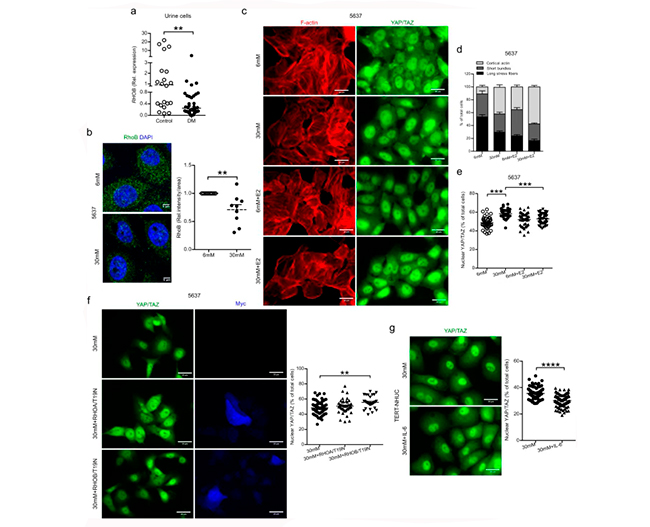
En células exfoliadas en orina de pacientes con diabetes, observaron una disminución de la expresión del ARNm de RHOB, así como también una disminución de la expresión del ARNm de RHOB y de la proteína en células uroepiteliales humanas expuestas a niveles altos de glucosa, 5637, como se muestra en la Figura 5 a y b. Sin embargo, el péptido psoriasina no indujo un efecto directo sobre la expresión del ARNm de RHOB en células tratadas con niveles altos de glucosa.
Curiosamente, el tratamiento con glucosa alta de células uroepiteliales humanas 5637 reveló menos fibras de estrés largas, más actina cortical, y predominantemente YAP/TAZ nuclear (Figuras 5c–e), un factor de transcripción responsable de la regulación negativa de la expresión de psoriasina. Por el contrario, en células normoglucémicas, YAP/TAZ se localizó tanto en el citoplasma como en el núcleo con forma celular normal y largas fibras de estrés. La misma observación se encontró en células uroepiteliales TERT-NHUC.
Para confirmar el papel de RhoB en la translocación de YAP/TAZ, las construcciones RHOB/T19N negativas dominantes y RHOA/T19N de control, se transfectaron por separado en células 5637 tratadas con alta glucosa seguido de la validación de YAP/TAZ nuclear. La expresión de RHOB/T19N resultó en un aumento de YAP/TAZ nuclear, mientras que no se observó ningún efecto con RHOA/T19N, lo que confirma el papel de RhoB negativo dominante en el aumento de la translocación de YAP/TAZ al núcleo (Figura 5f), sin efecto sobre el citoesqueleto de actina.
Además, el efecto de IL-6 en células TERT-NHUC cultivadas con alto contenido de glucosa resultó en una translocación de YAP/TAZ con menos YAP/TAZ nuclear (Figura 5g).
Y por otro lado, se investigó el efecto de RhoB sobre las comunidades bacterianas intracelulares (IBC) en vejigas de ratones infectados con E. coli., donde la reducción significativa de RhoB se asoció con un mayor número de IBC en la vejiga de ratones diabéticos, en comparación con la de ratones no diabéticos.
Efecto del estradiol sobre la carga bacteriana y depuración inducidas por niveles altos de glucosa
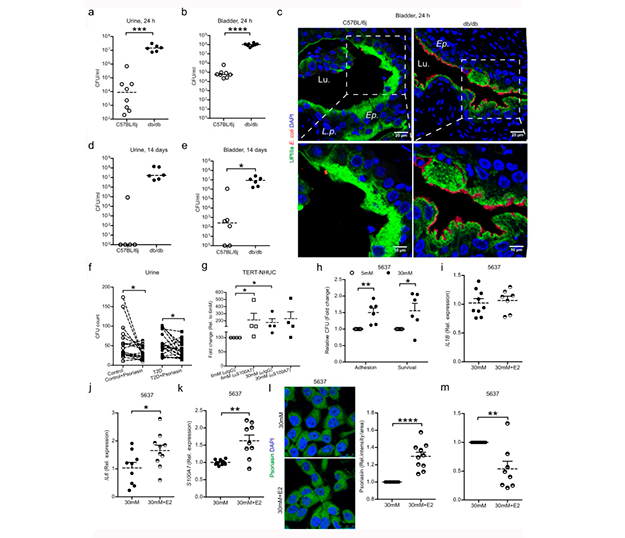
Para investigar el impacto de la glucosa en la infección y la depuración bacteriana, se infectaron ratones control diabéticos y no diabéticos con E. coli. Tras esto, observaron una mayor carga bacteriana en la orina y en la vejiga de los ratones diabéticos 24h después de la infección, como se muestra en la siguiente Figura 6 a y b.
Las células epiteliales superficiales de la vejiga de todos los ratones diabéticos presentaron numerosos CMI que ocupaban parcialmente la superficie epitelial y destruían el revestimiento celular, mientras que los ratones control no diabéticos presentaron un revestimiento celular intacto con escasos CMI, como se puede ver en la Figura 6c.
Una e incluso dos semanas después del inicio de la infección, los ratones diabéticos aún presentaban una mayor carga bacteriana en la orina y ≥106 bacterias en la vejiga (Figuras 6d y 6e), con numerosos CMI e inclusiones bacterianas fusionadas que ocupaban el epitelio, lo que indica una disminución de la depuración bacteriana.
Para confirmar la relevancia de la actividad de la psoriasina en la orina, se suplementaron muestras de controles y pacientes con diabetes tipo 2, con 5µM de péptido de psoriasina. Con esto, observaron una eliminación significativa de E. coli, tanto en las muestras de orina de control como en las de pacientes con diabetes tipo 2 (Figura 6f). Además, para confirmar la eliminación de E. coli mediada por la psoriasina, el bloqueo de la psoriasina con un anticuerpo monoclonal específico resultó en un número de bacterias dos veces mayor en el lisado de glucosa normal. Esto no se observó en los niveles altos de glucosa, lo que podría atribuirse en parte a los bajos niveles iniciales de psoriasina (Figura 6g).
Para dilucidar el impacto de la glucosa in vitro, infectaron con E. coli las células uroepiteliales 5637 (Figura 6h) y TERT-NHUC tratadas con niveles altos de glucosa. Esta infección mostró un aumento de la adhesión bacteriana y de la carga bacteriana intracelular, en comparación con concentraciones normales de glucosa.
Cabe destacar que el estradiol no influyó directamente en el ARNm de IL1B (Figura 6i), mientras que la expresión del ARNm de IL6 aumentó (Figura 6j), lo que indica que el estradiol actúa sobre IL-6, seguido de un aumento de S100A7 en los niveles de ARNm (Figura 6k) y proteína (Figura 6l). Además, se observó un aumento de la destrucción bacteriana, en comparación con las células uroepiteliales 5637 expuestas únicamente a niveles altos de glucosa (Figura 6m).
El tratamiento con estradiol de células uroepiteliales 5637 expuestas a altos niveles de glucosa inhibió la formación de fibras largas de estrés y haces cortos, y aumentó la formación de actina cortical después de 48h (Figuras 5c y 5 d anteriores). Además, el estradiol rescató las células tratadas con altos niveles de glucosa, localizando YAP/TAZ tanto en el citoplasma como en el núcleo (Figura 5e). Aunque el estradiol aumentó aún más la cantidad de actina cortical, el tratamiento resultó en una menor carga bacteriana (Figura 6m).
Para verificar el efecto de la red de actina sobre la carga bacteriana, se utilizó el inhibidor de actina Latrunculina B, lo que resultó en una carga bacteriana significativamente reducida, similar a la de las células tratadas con estradiol. Esto confirmó además que, a pesar del reordenamiento de la actina, el efecto sobre la carga bacteriana se vio influenciado por el efecto bactericida de la psoriasina inducida por estradiol.
Discusión y Conclusión del estudio de infección de orina derivada de la diabetes
En este estudio, se ha demostrado que los niveles elevados de glucosa comprometieron la respuesta inmunitaria innata y deterioraron la integridad epitelial de la vejiga. Estos hallazgos podrían explicar la observación clínica de que los pacientes con diabetes mal controlada presentan un mayor riesgo de infección de orina derivada de la diabetes recurrente, pielonefritis aguda y urosepticemia. Por lo tanto, se confirma que un control glucémico deficiente puede contribuir a un mayor riesgo de infecciones.
Los resultados demuestran que los niveles elevados de glucosa inhiben la psoriasina plasmática en individuos prediabéticos durante el pinzamiento hiperglucémico. Sin embargo, previamente, se han observado niveles séricos más bajos de AMP en pacientes diabéticos. Si bien se sabe que la insulina regula la expresión de la ARNasa a través de la vía PI3K, los resultados de este estudio de pinzamiento demuestran que la insulina no parece tener ningún efecto sobre la psoriasina. Por lo tanto, concluyen que la psoriasina está regulada por otra vía, lo cual se ve respaldado por la falta de influencia de PI3K en la expresión de psoriasina en células epiteliales mamarias, lo que confirma la influencia diferencial de la insulina en los AMP.
En consonancia con los hallazgos clínicos en pacientes con diabetes, también detectaron niveles más bajos de psoriasina en la vejiga urinaria de ratones diabéticos. No obstante, la infección por E. coli de células uroepiteliales tratadas con niveles altos de glucosa no logró aumentar la expresión de psoriasina. Asimismo, otros AMP, como la β-defensina-3 y la catelicidina, se vieron afectados en los queratinocitos tratados con niveles altos de glucosa y en los macrófagos, respectivamente. El efecto de la psoriasina es particularmente interesante, ya que se sugiere que es el AMP más potente y abundante contra E. coli. La ARNasa y la catelicidina LL-37 son diez veces menos potentes que la psoriasina, y esta última es, además, menos abundante en células epiteliales no estimuladas, lo que sugiere que son menos importantes en comparación con la psoriasina en la prevención de infecciones por E. coli. Y además de su efecto superior sobre E. coli, la psoriasina también tiene una alta potencia contra Enterococcus sp., lo que sugiere una función protectora más amplia contra las infecciones.
Por otro lado, la exfoliación de las células infectadas es un mecanismo común impulsado por el hospedador para eliminar la infección, pero también permite que las bacterias adheridas a las células vecinas infecten capas celulares más profundas. Un mayor riesgo de infección suele asociarse con la pérdida de la integridad de la barrera, lo que permite la invasión bacteriana. En este sentido, las proteínas de unión estrecha, distribuidas en las células epiteliales del tracto urinario, desempeñan un papel fundamental al proteger el tejido profundo de la invasión de patógenos.
Los hallazgos de este estudio revelan niveles más bajos de ocludina en individuos con prediabetes tras la infusión de glucosa, así como en pacientes con diabetes y en vejigas urinarias de ratones diabéticos. Estos hallazgos se asociaron con niveles más bajos de psoriasina. De igual manera, se ha descrito que los niveles elevados de glucosa reducen la expresión de ocludina en las células endoteliales de la retina humana y en la retina de ratones diabéticos. Aquí demostraron que las células uroepiteliales expuestas a niveles elevados de glucosa restauraron la expresión de ocludina tras el tratamiento con el péptido de psoriasina. Esto también concuerda con la expresión de ocludina mediada por psoriasina en los queratinocitos humanos, donde en pacientes no diabéticos, se ha descrito que otros AMP, como la catelicidina y la β-defensina-3, desencadenan la expresión de ocludina y claudinas en los queratinocitos humanos. El resultado de la restauración de la expresión del ARNm de OCLN en células uroepiteliales tratadas con alto contenido de glucosa y suplementadas con psoriasina, confirmó aún más el papel vital de la psoriasina en el mantenimiento de la integridad epitelial en la diabetes.
Por otra parte, en la diabetes, se forma un exceso de productos finales de la glicación avanzada y se acumula en los tejidos de la vejiga, lo que contribuye a la fijación de E. coli uropatógena (UPEC) y al inicio de la infección. Sin embargo, se ha demostrado un mayor número de sitios de unión que contienen manosa para las fimbrias de tipo 1 en la vejiga urinaria de ratones diabéticos. En el presente estudio, demostraron una regulación positiva de Mrc1 en ratones diabéticos infectados, pero no en ratones control, lo que facilita la colonización e invasión bacteriana, y respalda aún más la capacidad reducida para controlar las infecciones bacterianas.
Por su parte, el aumento de la expresión de caveolina 1 en la vejiga de ratones diabéticos crea una condición favorable para la entrada de bacterias. El resultado de este estudio destaca la importancia de la regulación negativa de la caveolina 1 mediada por psoriasina, que contribuye a una menor captación bacteriana en la célula. El reordenamiento de la caveolina 1 influye aún más en la estructura citoesquelética de la célula y en la organización de los filamentos de actina, y a su vez, facilita la capacidad de las bacterias para multiplicarse dentro de la célula. Además, la replicación bacteriana en capas celulares menos diferenciadas se ve inhibida por la red de actina más densa. Sin embargo, observaron que altas concentraciones de glucosa desencadenan la formación de más actina cortical, lo que podría potenciar el desarrollo de IBC, principalmente en las células paraguas terminalmente diferenciadas de ratones diabéticos.
La reorganización intracelular de la actina filamentosa resultó en la translocación de YAP/TAZ del citoplasma al núcleo. Esta reorganización también está regulada por la caveolina 1. La expresión de psoriasina depende de la densidad y la morfología celular, y YAP/TAZ nuclear regula significativamente la expresión de psoriasina en el carcinoma de células escamosas. Por lo tanto, la observación del aumento de la expresión de caveolina 1 mediada por glucosa, la formación de actina cortical y la translocación de YAP/TAZ al núcleo, enfatizó una vía mecanicista de regulación negativa de la psoriasina, confirmada en dos células uroepiteliales diferentes: 5637 y TERT-NHUC.
Asimismo, la familia Rho de GTPasas pequeñas es un regulador clave de los reordenamientos del citoesqueleto de actina y se asocia con una mayor formación de IBC en células epiteliales de vejiga humana, 5637. Los resultados de este estudio respaldan los hallazgos de la regulación negativa de RhoB mediada por altos niveles de glucosa y el aumento de la carga bacteriana intracelular en ratones diabéticos. Además, demostraron que RhoB influye en la translocación de YAP/TAZ nuclear.
Por otro lado, el aumento de la carga bacteriana y el exceso de IBC detectados en las vejigas de ratones diabéticos, incluso después de 14 días de infección por E. coli, indican una incapacidad para restringir el crecimiento y la distribución bacteriana en comparación con los controles no diabéticos. Además, las bacterias que residen en el tejido más profundo pueden servir como reservorios y contribuir a futuras infecciones. Asimismo, demostraron que los cambios mediados por la alta glucosa en la expresión de psoriasina, ocludina, Mrc1 y caveolina 1, resultaron en un aumento de la carga bacteriana. Por lo tanto, este resultado contribuye a la comprensión de la mayor susceptibilidad a UPEC en modelos de ratones prediabéticos y diabéticos y la disminución de la depuración de UPEC en ratones diabéticos.
Recientemente, se ha demostrado que el estradiol aumenta la tolerancia a la glucosa y la sensibilidad a la insulina en ratones ovariectomizados con agotamiento de estrógenos. En este estudio, demostraron el efecto directo del estradiol sobre la expresión de IL-6, un posible mecanismo para una vía de psoriasina mediada por estradiol sin la participación de IL-1B en células uroepiteliales humanas tratadas con niveles altos de glucosa. Además, demostraron que el estradiol resultó en un aumento de la destrucción bacteriana de las células uroepiteliales infectadas con E. coli, al translocar YAP/TAZ de vuelta al citoplasma y restaurar la expresión de psoriasina en células uroepiteliales tratadas con niveles altos de glucosa sin afectar el citoesqueleto. También se ha demostrado que la señalización del receptor de estrógeno afecta la expresión de psoriasina. Por lo tanto, se cree que el estradiol influye en la regulación de la respuesta inmunitaria. Además, sus observaciones confirman el efecto beneficioso del estrógeno también en condiciones diabéticas y de hiperglucemia.
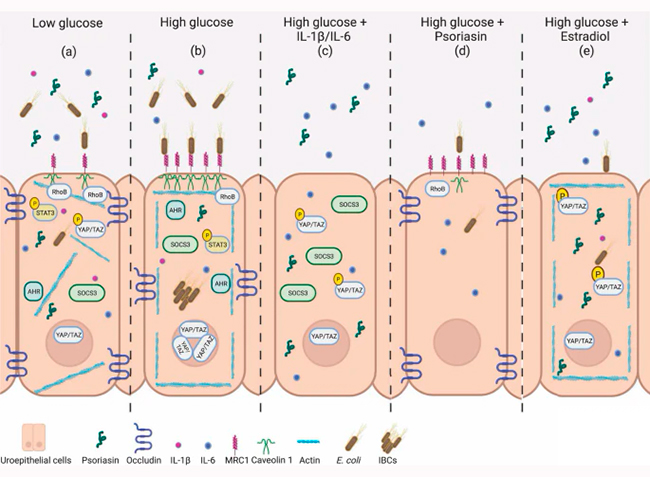
En conclusión, la hiperglucemia compromete la psoriasina a través de las vías IL-6 y YAP/TAZ, que afectan la barrera epitelial y causan alteraciones en la membrana celular. Estos cambios, crean además un entorno favorable para la infección bacteriana. Los resultados de este estudio sugieren que la psoriasina, junto con otros factores, busca contrarrestar las consecuencias negativas de la hiperglucemia. La hipótesis de los investigadores es que estos factores, si bien actúan de forma parcialmente independiente, podrían estar interrelacionados, lo que potencia su efecto. En conjunto, los datos sugieren que la psoriasina es un péptido antimicrobiano importante en la eliminación de bacterias del tracto urinario en la diabetes y podría, en el futuro, servir como posible diana para nuevos fármacos terapéuticos.
Fuente de este artículo
Este artículo fue publicado originalmente en la revista Nature Communications bajo la referencia: Mohanty, S., Kamolvit, W., Scheffschick, A. et al. Diabetes downregulates the antimicrobial peptide psoriasin and increases E. coli burden in the urinary bladder. Nat Commun 13, 4983 (2022). https://doi.org/10.1038/s41467-022-32636-y; estando disponible con acceso libre en la propia web de Nature Communications.